La capacidad de ver el mundo que nos rodea es una de las experiencias más ricas y complejas que poseemos. Pero la visión no es un simple acto de 'mirar'; es un proceso activamente construido por nuestro cerebro. Aunque los ojos captan la luz y forman imágenes iniciales, es en las intrincadas redes neuronales de la corteza visual donde estas señales se transforman en la percepción consciente que experimentamos: rostros, objetos, colores, movimientos y la profundidad del espacio. Este viaje comienza en áreas cerebrales específicas, cada una con roles definidos pero interconectadas de manera compleja para dar sentido al torrente de información visual.

La Corteza Visual Primaria (V1)
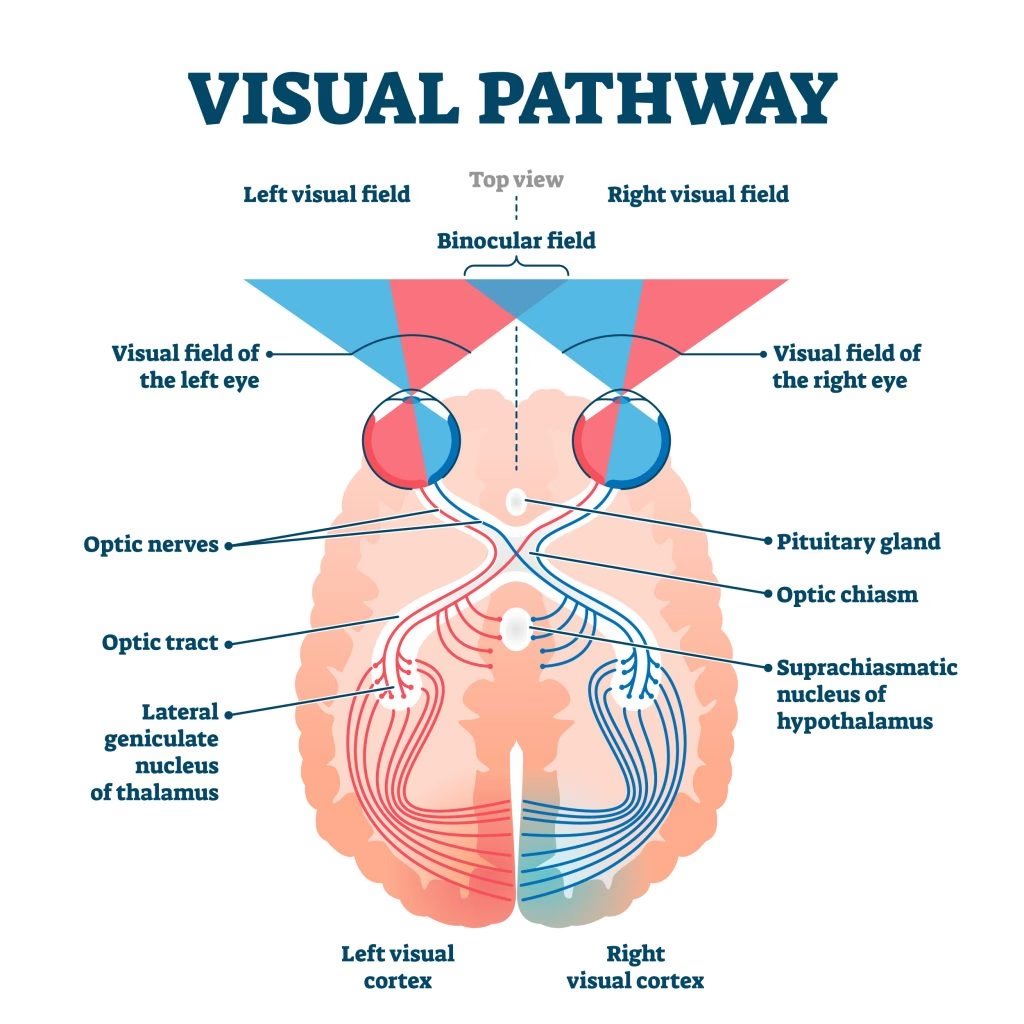
El primer punto de procesamiento cortical para la información visual es la corteza visual primaria, conocida como V1. Esta área se localiza en y alrededor de la cisura calcarina, en el lóbulo occipital, en la parte posterior de nuestro cerebro. Cada hemisferio cerebral contiene una V1 que recibe información directamente del núcleo geniculado lateral ipsilateral, el cual, a su vez, recibe señales del hemicampo visual contralateral. Esto significa que la V1 derecha procesa información del lado izquierdo del campo visual, y viceversa.
Las neuronas en la corteza visual, incluida V1, se activan cuando aparecen estímulos visuales dentro de su campo receptivo. Por definición, el campo receptivo es la región del campo visual total que provoca que una neurona dispare un potencial de acción. Sin embargo, una neurona puede responder mejor a un subconjunto específico de estímulos dentro de su campo receptivo. Esta propiedad se denomina neuronal tuning. En las áreas visuales tempranas, como V1, las neuronas tienen una sintonización más simple. Por ejemplo, una neurona en V1 puede disparar en respuesta a cualquier estímulo vertical dentro de su campo receptivo.
Una característica fundamental de V1 es su organización retinotópica. Esto significa que las células vecinas en V1 tienen campos receptivos que corresponden a porciones adyacentes del campo visual. Esta organización espacial permite una representación sistemática del mundo visual dentro de V1. Además, estudios recientes han explorado el papel de la modulación contextual en V1, donde la percepción de un estímulo no solo está influenciada por el estímulo en sí, sino también por el contexto circundante. Esto subraya las complejas capacidades de procesamiento de V1 para moldear nuestras experiencias visuales.
La corteza visual recibe su suministro de sangre principalmente de la rama calcarina de la arteria cerebral posterior. Es interesante notar que el tamaño de V1, V2 y V3 puede variar hasta tres veces entre individuos, una diferencia que es parcialmente heredada.
El Área Visual Secundaria (V2)
Inmediatamente después de V1 en la jerarquía de procesamiento se encuentra el área visual secundaria, o V2, también conocida como corteza preestriada. V2 recibe fuertes conexiones de avance desde V1 (directamente y a través del pulvinar) y envía conexiones robustas a V3, V4 y V5. Además, desempeña un papel crucial en la integración y procesamiento de la información visual.
Las conexiones de avance de V1 a V2 contribuyen al procesamiento jerárquico de los estímulos visuales. Las neuronas en V2 se basan en las características básicas detectadas en V1, extrayendo atributos visuales más complejos como textura, profundidad y color. Este procesamiento jerárquico es esencial para la construcción de una representación más matizada y detallada de la escena visual.
Asimismo, las conexiones recíprocas de retroalimentación de V2 a V1 desempeñan un papel significativo en la modulación de la actividad de las neuronas de V1. Se cree que este bucle de retroalimentación está involucrado en procesos como la atención, el agrupamiento perceptual y la segregación figura-fondo. La interacción dinámica entre V1 y V2 destaca la naturaleza intrincada del procesamiento de información dentro del sistema visual.
Las conexiones de V2 con áreas visuales posteriores, incluidas V3, V4 y V5, contribuyen a la formación de una red distribuida para el procesamiento visual. Estas conexiones permiten la integración de diferentes características visuales, como el movimiento y la forma, a través de múltiples etapas de la jerarquía visual. Anatómicamente, V2 se divide en cuatro cuadrantes: una representación dorsal y una ventral en los hemisferios izquierdo y derecho. Juntas, estas cuatro regiones proporcionan un mapa completo del mundo visual.
V2 comparte muchas propiedades con V1: las células están sintonizadas a propiedades simples como la orientación, la frecuencia espacial y el color. Sin embargo, las respuestas de muchas neuronas de V2 también están moduladas por propiedades más complejas, como la orientación de contornos ilusorios, la disparidad binocular, y si el estímulo forma parte de la figura o del fondo. Investigaciones recientes han demostrado que las células de V2 muestran una pequeña cantidad de modulación atencional (más que V1, menos que V4), están sintonizadas a patrones moderadamente complejos y pueden ser activadas por múltiples orientaciones en diferentes subregiones dentro de un solo campo receptivo.
Se argumenta que toda la vía visual ventral hacia el hipocampo es importante para la memoria visual. Esta teoría, a diferencia de la dominante, predice que las alteraciones en la memoria de reconocimiento de objetos podrían resultar de la manipulación en V2, un área altamente interconectada dentro de la vía ventral de las cortezas visuales. En el cerebro del mono, esta área recibe fuertes conexiones de avance de la corteza visual primaria (V1) y envía fuertes proyecciones a otras cortezas visuales secundarias (V3, V4 y V5).
La mayoría de las neuronas de esta área en primates están sintonizadas a características visuales simples como la orientación, la frecuencia espacial, el tamaño, el color y la forma. Estudios anatómicos implican a la capa 3 del área V2 en el procesamiento de información visual. En contraste con la capa 3, la capa 6 de la corteza visual está compuesta por muchos tipos de neuronas, y su respuesta a los estímulos visuales es más compleja. En un estudio, se encontró que las células de la Capa 6 de la corteza V2 desempeñan un papel muy importante en el almacenamiento de la Memoria de Reconocimiento de Objetos, así como en la conversión de memorias de objetos a corto plazo en memorias a largo plazo.
El Tercer Complejo Visual (V3) y Sus Subdivisiones
El término tercer complejo visual se refiere a la región de la corteza ubicada inmediatamente delante de V2, que incluye la región denominada área visual V3 en humanos. La nomenclatura de "complejo" se justifica porque aún existe cierta controversia sobre la extensión exacta del área V3, y algunos investigadores proponen que la corteza ubicada delante de V2 puede incluir dos o tres subdivisiones funcionales.
Por ejemplo, David Van Essen y otros (1986) han propuesto la existencia de una "V3 dorsal" en la parte superior del hemisferio cerebral, que es distinta de la "V3 ventral" (o área posterior ventral, VP) ubicada en la parte inferior del cerebro. La V3 dorsal y ventral tienen conexiones distintas con otras partes del cerebro, parecen diferentes en secciones teñidas con una variedad de métodos, y contienen neuronas que responden a diferentes combinaciones de estímulo visual (por ejemplo, las neuronas selectivas al color son más comunes en la V3 ventral).
También se han reportado subdivisiones adicionales, incluidas V3A y V3B, en humanos. Estas subdivisiones se encuentran cerca de la V3 dorsal, pero no colindan con V2. La V3 dorsal se considera normalmente parte de la vía dorsal, recibiendo entradas de V2 y del área visual primaria y proyectando a la corteza parietal posterior. Anatómicamente, podría estar ubicada en el área de Brodmann 19. Braddick, utilizando fMRI, ha sugerido que el área V3/V3A puede desempeñar un papel en el procesamiento del movimiento global.
Otros estudios prefieren considerar la V3 dorsal como parte de un área más grande, denominada área dorsomedial (DM), que contiene una representación de todo el campo visual. Las neuronas en el área DM responden al movimiento coherente de patrones grandes que cubren porciones extensas del campo visual. La V3 ventral (VP) tiene conexiones mucho más débiles de la corteza visual primaria y conexiones más fuertes con la corteza temporal inferior. Si bien estudios anteriores proponían que VP contenía una representación solo de la parte superior del campo visual (por encima del punto de fijación), trabajos más recientes indican que esta área es más extensa de lo que se creía, y al igual que otras áreas visuales, puede contener una representación visual completa. La VP revisada y más extensa es referida como el área posterior ventrolateral (VLP) por Rosa y Tweedale.
El Área Visual del Lóbulo Temporal Medio (V5 o MT)
El área visual del lóbulo temporal medio (MT o V5) es una región de la corteza visual extraestriada. En varias especies de monos del Nuevo y Viejo Mundo, el área MT contiene una alta concentración de neuronas selectivas a la dirección del movimiento. Se cree que el MT en primates desempeña un papel importante en la percepción del movimiento, la integración de señales de movimiento locales en perceptos globales y la guía de algunos movimientos oculares.
El MT está conectado a una amplia gama de áreas cerebrales corticales y subcorticales. Recibe entradas de las áreas corticales visuales V1, V2 y V3 dorsal (área dorsomedial), las regiones koniocelulares del LGN y el pulvinar inferior. El patrón de proyecciones a MT cambia ligeramente entre las representaciones de los campos visuales foveal y periférico, recibiendo este último entradas de áreas ubicadas en la corteza de la línea media y la región retrosplenial. Una visión estándar es que V1 proporciona la entrada "más importante" a MT. Sin embargo, varios estudios han demostrado que las neuronas en MT son capaces de responder a información visual, a menudo de manera selectiva a la dirección, incluso después de que V1 ha sido destruida o inactivada. Además, investigaciones de Semir Zeki y colaboradores han sugerido que ciertos tipos de información visual pueden llegar a MT incluso antes de que llegue a V1.
MT envía su principal salida a las áreas ubicadas en la corteza inmediatamente circundante, incluidas las áreas FST, MST y V4t (media luna temporal media). Otras proyecciones de MT se dirigen a las áreas relacionadas con los movimientos oculares de los lóbulos frontal y parietal (campo ocular frontal y área intraparietal lateral).
Los primeros estudios de las propiedades electrofisiológicas de las neuronas en MT mostraron que una gran parte de las células están sintonizadas a la velocidad y dirección de los estímulos visuales en movimiento. Los estudios de lesión también han apoyado el papel de MT en la percepción del movimiento y los movimientos oculares. Estudios neuropsicológicos de un paciente incapaz de ver movimiento, viendo el mundo en una serie de 'fotogramas' estáticos en su lugar, sugirieron que V5 en primates es homólogo a MT en humanos. Sin embargo, dado que las neuronas en V1 también están sintonizadas a la dirección y velocidad del movimiento, estos resultados iniciales dejaron abierta la pregunta de qué podía hacer MT que V1 no pudiera.
Se ha realizado mucho trabajo en esta región, ya que parece integrar señales de movimiento visual locales en el movimiento global de objetos complejos. Por ejemplo, la lesión de V5 provoca déficits en la percepción del movimiento y el procesamiento de estímulos complejos. Contiene muchas neuronas selectivas para el movimiento de características visuales complejas (extremos de líneas, esquinas). La microestimulación de una neurona ubicada en V5 afecta la percepción del movimiento. Por ejemplo, si se encuentra una neurona con preferencia por el movimiento hacia arriba en el V5 de un mono y se estimula con un electrodo, es más probable que el mono reporte movimiento 'hacia arriba' cuando se le presentan estímulos que contienen componentes 'izquierda' y 'derecha' además de 'hacia arriba'. Todavía existe mucha controversia sobre la forma exacta de los cálculos realizados en el área MT, y algunas investigaciones sugieren que el movimiento de características de hecho ya está disponible en niveles inferiores del sistema visual como V1.
El MT se organiza en columnas de dirección. DeAngelis argumentó que las neuronas de MT también se organizaban en función de su sintonización a la disparidad binocular.
Comparativa de Áreas Visuales Clave
| Área | Ubicación (relativa a V1) | Entradas Principales | Salidas Principales | Rol Primario | Complejidad de Sintonización |
|---|---|---|---|---|---|
| V1 | Cisura Calcarina (Lóbulo Occipital) | LGN | V2, Pulvinar | Procesamiento inicial, detección de bordes, orientación, frecuencia espacial | Simple (orientación, frecuencia) |
| V2 | Inmediatamente delante de V1 | V1, Pulvinar | V3, V4, V5 | Integración, textura, profundidad, color, contornos ilusorios, figura-fondo | Moderada (combina características de V1) |
| V3 Dorsal (DM) | Delante de V2 (parte superior) | V1, V2 | Corteza Parietal Posterior | Movimiento global, parte de la vía dorsal ('dónde'/'cómo') | Responde a movimiento coherente |
| V3 Ventral (VP/VLP) | Delante de V2 (parte inferior) | V2 (débil de V1) | Corteza Temporal Inferior | Color (más común en V3v), parte de la vía ventral ('qué') | Varía, algunos selectivos al color |
| V5 (MT) | Corteza Extraestriada (Lóbulo Temporal Medio) | V1, V2, V3 Dorsal (DM), LGN, Pulvinar Inferior | FST, MST, V4t, Áreas de Movimiento Ocular (Frontal/Parietal) | Percepción y procesamiento del movimiento, integración local-global | Alta (velocidad, dirección, movimiento complejo) |
Preguntas Frecuentes sobre la Percepción Visual y el Cerebro
¿Qué es el campo receptivo de una neurona visual?
Es la región específica dentro de todo el campo visual que, al ser estimulada, provoca que esa neurona dispare un potencial de acción. Es la 'área de vigilancia' de la neurona.
¿Qué significa que V1 tiene una organización retinotópica?
Significa que las células vecinas en V1 procesan información de partes adyacentes del campo visual. Esto crea un mapa espacial organizado del mundo visual en V1, similar a cómo se proyecta en la retina.
¿Cuál es la diferencia principal en la función entre V1 y V2?
V1 realiza un procesamiento inicial de características simples como la orientación y la frecuencia espacial. V2 se basa en esta información para procesar características más complejas como textura, profundidad, color, contornos ilusorios y si algo es figura o fondo, e integra información de V1 de forma más compleja.
¿Las áreas visuales solo procesan información en una dirección (de V1 hacia V5)?
No, aunque existe una jerarquía de procesamiento (flujo de avance), también hay importantes conexiones de retroalimentación (por ejemplo, de V2 a V1) que modulan la actividad y están implicadas en procesos como la atención y el agrupamiento perceptual.
¿Qué área cerebral es crucial para la percepción del movimiento?
El área visual del lóbulo temporal medio, conocida como V5 o MT, es fundamental para la percepción del movimiento, la integración de señales de movimiento locales y la guía de movimientos oculares. Las neuronas en esta área son altamente selectivas a la dirección y velocidad del movimiento.
La visión es, en esencia, un acto de construcción cerebral. Desde la detección inicial de características básicas en V1 hasta el procesamiento de movimiento complejo en V5, la integración de información en V2 y la especialización en las subdivisiones de V3, una red compleja de áreas visuales trabaja en concierto. Cada área contribuye con un tipo específico de procesamiento, pero es la interacción dinámica entre ellas, a través de vías de avance y retroalimentación, lo que nos permite percibir el mundo visual de manera coherente y significativa. Este intrincado sistema neuronal es un testimonio de la asombrosa capacidad del cerebro humano para transformar la luz en experiencia.
Si quieres conocer otros artículos parecidos a El Cerebro y la Visión: Un Viaje Neuronal puedes visitar la categoría Neurociencia.