En el complejo entramado de nuestro cerebro, existen regiones altamente especializadas dedicadas a tareas específicas. Una de las funciones visuales más cruciales es la percepción del movimiento, una habilidad esencial para interactuar con nuestro entorno, desde esquivar un obstáculo hasta atrapar un objeto en movimiento. Esta proeza perceptiva depende en gran medida de un área particular en la corteza cerebral: el área temporal medial, conocida comúnmente como área MT.

- ¿Qué es el Área MT?
- MT y MST: El Equipo Dinámico
- La Ruta del Movimiento: Desde la Retina hasta MT
- MT: Un Nodo Crucial en el Desarrollo Visual
- Más Allá de la Detección Simple: Células Patrón y Componente
- El Problema de la Apertura y su Solución en MT
- Modelos Computacionales y la Selectividad de MT
- Comparativa de Células MT
- Comparativa de Entradas V1 a MT
- Preguntas Frecuentes sobre el Área MT
¿Qué es el Área MT?
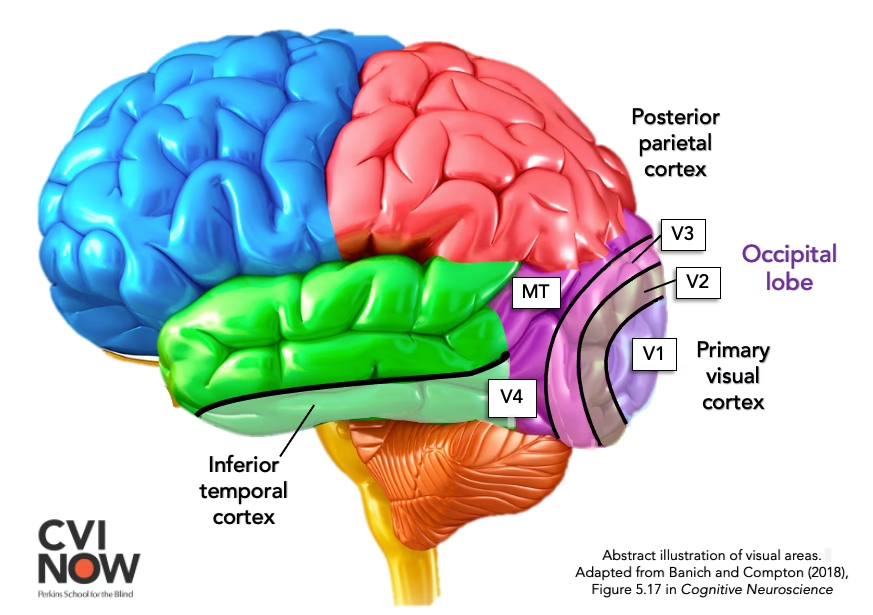
El área MT, o área temporal medial, es una región especializada dentro de la corteza visual extrastriada. Su nombre completo es área temporal medial, y a menudo se la conoce también como V5. Esta área se distingue por su papel fundamental en la detección y el procesamiento del movimiento visual. Anatómicamente, el área MT se localiza justo delante de la unión entre el lóbulo temporal y el lóbulo occipital en la parte posterior del cerebro.
El área MT no trabaja de forma aislada; forma parte esencial de una vía visual más amplia conocida como la corriente dorsal. Esta vía, en contraste con la corriente ventral (más asociada con la identificación de objetos y formas), se encarga principalmente de procesar la ubicación espacial, el movimiento y la orientación de los objetos. Piensa en la diferencia entre reconocer que algo es una pelota (corriente ventral) y saber dónde está y cómo se mueve (corriente dorsal, con MT como actor clave).
MT y MST: El Equipo Dinámico
Estrechamente relacionada con el área MT se encuentra el área temporal superior medial (MST). Ambas áreas, MT y MST, trabajan en conjunto para el procesamiento del movimiento. La información visual relacionada con la dirección del movimiento llega primero a MT, que recibe la entrada de neuronas selectivas a la dirección en el área visual primaria (V1). Mientras que las neuronas de V1 pueden responder a movimientos locales o bordes, las neuronas en MT son expertas en detectar el movimiento coherente dentro de parches o regiones del campo visual.
Una vez que MT ha procesado este movimiento coherente, envía esta información al área MST. MST integra esta información de movimiento coherente de áreas más amplias de la escena visual, lo que es crucial para detectar el flujo óptico. El flujo óptico es el patrón aparente de movimiento de los objetos, superficies y bordes en una escena visual causado por el movimiento relativo entre el observador y la escena. Esto es vital, por ejemplo, para navegar por el entorno o para realizar acciones complejas como atrapar un objeto en movimiento, donde MT y MST nos ayudan a seguir la trayectoria y ajustar nuestros movimientos.
La importancia de MT y MST en el procesamiento del movimiento ha sido respaldada por diversas investigaciones. Por ejemplo, la estimulación eléctrica directa del área MT puede influir significativamente en nuestra capacidad para percibir y procesar el movimiento visual, demostrando su papel causal en esta función.
La Ruta del Movimiento: Desde la Retina hasta MT
Se cree que la "vía del movimiento" es una corriente específica dentro del sistema visual que se especializa en la representación de señales de movimiento. Esta vía comienza en las células parasol de la retina, que son particularmente sensibles al contraste y al movimiento. La información viaja luego a través de diversas estructuras subcorticales y llega a la corteza visual primaria (V1).
En V1, existen neuronas que son selectivas a la dirección. Esto significa que responden fuertemente cuando un objeto o un campo de puntos aleatorios se mueve en una dirección específica, pero muestran poca respuesta a otras direcciones. El área MT es conocida por ser la primera estación cortical importante que recibe estas entradas direccionalmente selectivas de V1. Las neuronas en MT integran estas señales de movimiento locales de V1 para detectar patrones de movimiento más amplios y coherentes. Esta integración es un paso esencial para comprender el movimiento de objetos completos en lugar de solo el movimiento de sus bordes.
MT: Un Nodo Crucial en el Desarrollo Visual
Investigaciones recientes han puesto de manifiesto que el área MT no solo es fundamental para la percepción del movimiento en la edad adulta, sino que también desempeña un papel central en el desarrollo del sistema visual y en el establecimiento de comportamientos visuomotores. Un estudio relevante utilizó un modelo de lesión unilateral del área MT en crías de tití antes de examinar las consecuencias anatómicas, fisiológicas y conductuales en la edad adulta.
Los resultados fueron impactantes. En los animales adultos que habían sufrido la lesión temprana en MT, se observaron alteraciones significativas en comportamientos visuomotores orientados a objetivos, como alcanzar y agarrar. Específicamente, mostraron perturbaciones en la precisión de la secuencia de alcance y agarre, tanto para objetos estáticos como en movimiento. Curiosamente, en la tarea de objetos en movimiento, los animales lesionados tendían a iniciar el alcance de forma prematura y mostraban una mayor magnitud de error en el agarre fallido.
Más allá de los déficits conductuales, el estudio reveló cambios anatómicos y fisiológicos duraderos. Se encontraron alteraciones en la citoarquitectura de áreas de la corriente dorsal asociadas con el alcance (áreas parietal y medial intraparietal) y el agarre (área intraparietal anterior). Además, se observó un impacto significativo en la anatomía del área V1 y en la selectividad direccional de las neuronas de V1 en la zona de proyección de la lesión.
Estos hallazgos sugieren que el área MT actúa como un nodo crucial en el desarrollo del sistema visual de los primates. Su maduración temprana, que parece ocurrir en paralelo con la corteza visual primaria (V1) e incluso podría ser independiente de la entrada de V1 en las primeras etapas, le permitiría servir como un "ancla" para el establecimiento de la red de la corriente dorsal. La pérdida temprana de este nodo central perturba la arquitectura y función de áreas tanto 'aguas arriba' (como V1) como 'aguas abajo' en la vía dorsal, con consecuencias permanentes en los comportamientos guiados visualmente.
El fenómeno del 'blindsight', donde individuos con daño en V1 aún pueden realizar ciertas tareas visuales (como detectar movimiento) sin tener conciencia visual, también ha implicado a la corriente dorsal y al área MT como sustrato neural. Estudios en pacientes con blindsight han demostrado que MT permanece activo cuando se presentan estímulos visuales dentro de su escotoma (el área ciega causada por el daño en V1), reforzando la idea de que MT puede procesar movimiento incluso con una entrada V1 comprometida.
Más Allá de la Detección Simple: Células Patrón y Componente
Aunque la función principal de MT es procesar movimiento, las neuronas dentro de esta área no responden de una manera uniforme. Un hallazgo clave, basado en la estimulación con patrones de rayas cruzadas (conocidos como "plaids"), ha llevado a dividir las neuronas de MT en dos tipos principales: células "patrón" y células "componente".
Cuando se presenta un "plaid" (formado por la superposición de dos rejillas que se mueven en direcciones diferentes), los humanos percibimos un movimiento global del patrón en una dirección diferente a la de las rejillas individuales. Por ejemplo, dos rejillas que se mueven horizontalmente en direcciones opuestas pero con orientaciones inclinadas pueden percibirse como moviéndose verticalmente.
- Células Componente: Aproximadamente un tercio de las neuronas de MT se comportan como células componente. Estas células no responden al movimiento global del patrón, sino a la dirección del movimiento de las rejillas individuales que componen el patrón. Su curva de sintonización direccional mostrará dos picos, uno por cada dirección de componente.
- Células Patrón: Otro tercio de las neuronas de MT son células patrón. Estas sí son selectivas a la dirección del movimiento global del patrón. Su curva de sintonización direccional tendrá un solo pico, centrado en la dirección del movimiento percibido del patrón completo.
- El tercio restante muestra propiedades intermedias.
La existencia de estos dos tipos de células plantea la pregunta de cómo el cerebro pasa de codificar el movimiento de los componentes individuales a codificar el movimiento del patrón global. La teoría predominante sugería una jerarquía donde las células patrón sumaban las salidas de las células componente. Sin embargo, modelos computacionales más recientes proponen un mecanismo alternativo.
El Problema de la Apertura y su Solución en MT
Un desafío fundamental en la percepción del movimiento es el "problema de la apertura". Las neuronas visuales en áreas tempranas como V1 tienen campos receptivos pequeños. Cuando un borde largo y orientado se mueve a través de estos pequeños campos receptivos (la "apertura"), la neurona solo puede "ver" el movimiento del borde perpendicular a su orientación, no la dirección real del movimiento del objeto completo. Por ejemplo, si un lápiz horizontal se mueve diagonalmente hacia arriba y a la derecha, una neurona V1 con un pequeño campo receptivo sobre el borde del lápiz solo podría detectar un movimiento vertical hacia arriba.
El área MT juega un papel crucial en resolver este problema integrando información de movimiento de diferentes partes del campo visual. Las neuronas de MT, con campos receptivos más grandes que las de V1, pueden recibir información de múltiples neuronas de V1 que cubren diferentes partes de un objeto en movimiento.
Además, el texto sugiere que la solución al problema de la apertura en MT no solo proviene de la integración de señales de neuronas V1 "estándar" (complejas), sino también de la interacción con otros tipos de neuronas V1 especializadas:
- Neuronas V1 "End-stopped": Estas neuronas responden fuertemente a los "terminadores" (extremos) de las líneas o a los puntos de cruce. Los terminadores proporcionan información de movimiento menos ambigua que los bordes largos. Estas neuronas son cruciales para diferenciar la información de movimiento ambigua de los bordes de la información no ambigua de los terminadores.
- Neuronas V1 con Campos Receptivos Extra-Clásicos (V1-ECRF): Estas neuronas son sensibles a la forma y tienen entornos supresores sensibles al contraste. Su actividad se suprime en los puntos de cruce (terminadores extrínsecos) donde el entorno supresor está altamente activado. Esto permite que la información de los terminadores intrínsecos (extremos reales de los objetos) tenga más influencia.
La combinación de las señales de estos diferentes tipos de neuronas V1 (complejas, end-stopped, ECRF) en MT ayuda al cerebro a determinar la dirección de movimiento real de un objeto, superando la ambigüedad local del problema de la apertura.
Modelos Computacionales y la Selectividad de MT
Los modelos computacionales han sido herramientas valiosas para entender cómo el área MT genera la selectividad a patrones y componentes. Un modelo propuesto sugiere que la sintonización de las neuronas de MT (si responden a patrones o componentes) depende de la fuerza relativa de las conexiones que reciben de los tres tipos de neuronas V1 mencionados (complejas, end-stopped, V1-ECRF).
En este modelo, la dominancia de la entrada de las neuronas V1 "end-stopped" (que responden a terminadores) conduce a la selectividad a patrones en MT. Esto se debe a que los puntos de cruce (terminadores extrínsecos) en un "plaid" representan la dirección del movimiento del patrón. La propagación de la actividad de estas neuronas en MT permitiría detectar el movimiento global.
Por otro lado, la dominancia de la entrada de las neuronas V1-ECRF (con entornos supresores) resultaría en la selectividad a componentes. Estas neuronas suprimen la actividad en los puntos de cruce, reduciendo su influencia en la percepción del movimiento global y favoreciendo la respuesta a los movimientos individuales de los bordes.
Este modelo también explica otras características observadas en las neuronas de MT:
- Dependencia del Contraste: La selectividad a patrones en MT disminuye cuando las rejillas que forman el "plaid" tienen contrastes diferentes. El modelo lo explica porque la actividad de las neuronas V1-ECRF (que influyen en la selectividad a patrones) depende del contraste del estímulo.
- Dinámica Temporal: La detección del movimiento del patrón en MT muestra un retraso temporal (de 50 a 70 ms) en comparación con la detección del movimiento del componente. Esto se explica en el modelo por el tiempo que tarda la información de movimiento en propagarse desde los terminadores (puntos de cruce) a través de la red neuronal de MT para cubrir todo el estímulo. La detección del componente, al basarse en la actividad local a lo largo de los bordes, no requiere esta propagación extensa y, por lo tanto, es más rápida.
Estos hallazgos sugieren que la detección de movimiento de patrón en MT no es simplemente la suma de las señales de componente, sino que depende críticamente de la detección e integración de la información de los terminadores, especialmente los extrínsecos, que son puntos clave para resolver la ambigüedad del movimiento.
Comparativa de Células MT
| Característica | Células Patrón | Células Componente |
|---|---|---|
| Respuesta a "Plaids" | Movimiento global del patrón | Movimiento de las rejillas individuales |
| Curva de Sintonización Direccional | Un solo pico (dirección del patrón) | Dos picos (direcciones de los componentes) |
| Dependencia del Contraste | Alta, disminuye con contrastes diferentes | Baja, relativamente invariante al contraste |
| Dinámica Temporal | Muestra un retraso (propagación de señal) | Respuesta rápida (basada en bordes locales) |
| Influencia de Terminadores Extrínsecos | Alta (clave para la sintonización) | Baja (suprimida por V1-ECRF) |
Comparativa de Entradas V1 a MT
| Tipo de Neurona V1 | Función Principal | Respuesta a Estímulos con Bordes/Terminadores | Influencia en Neuronas MT (según modelo) |
|---|---|---|---|
| Compleja Estándar | Detección de movimiento local (sujeto a problema de apertura) | Alta actividad en bordes y terminadores (ambigua) | Entrada básica de movimiento a MT |
| End-Stopped | Detección de terminadores (extremos, cruces) | Alta actividad en terminadores (menos ambigua) | Inhibe neuronas MT de segmentación; clave para sintonización a patrón |
| ECRF (Extra-Classical Receptive Field) | Detección de forma, entornos supresores | Actividad suprimida en terminadores extrínsecos (cruces) | Excita neuronas MT de integración; clave para sintonización a componente (al suprimir cruces) |
Preguntas Frecuentes sobre el Área MT
¿Qué significa MT en neurociencia?
MT significa Área Temporal Medial (Middle Temporal area). A veces también se le llama V5.
¿Cuál es la función principal del Área MT?
La función principal del Área MT es la detección y el procesamiento del movimiento visual. Es crucial para percibir cómo se mueven los objetos en nuestro entorno.
¿El Área MT forma parte de la corriente visual dorsal o ventral?
El Área MT es una parte esencial de la corriente visual dorsal, que está implicada en el procesamiento de la ubicación espacial, el movimiento y la orientación de los objetos.
¿Qué son las células patrón y componente en MT?
Son dos tipos de neuronas en MT que responden de manera diferente a patrones de movimiento complejos como los "plaids". Las células componente responden al movimiento de los elementos individuales, mientras que las células patrón responden al movimiento global del patrón completo.
¿Qué sucede si el Área MT sufre daño en la infancia?
Estudios en modelos animales sugieren que el daño temprano en MT puede tener consecuencias duraderas en el desarrollo de la corriente dorsal, afectar la función de V1 y alterar significativamente los comportamientos visuomotores, como alcanzar y agarrar objetos.
¿Cómo ayuda el Área MT a resolver el problema de la apertura?
MT integra información de neuronas V1 que responden a diferentes partes de un objeto en movimiento. Además, utiliza información de neuronas V1 especializadas (como las end-stopped y ECRF) que responden a terminadores (extremos y cruces), los cuales proporcionan información de movimiento menos ambigua que los bordes largos.
En resumen, el área MT es una estación de procesamiento de movimiento de vital importancia en el cerebro, no solo por su papel en la percepción adulta, sino también por su influencia fundamental en el cableado y la función del sistema visual durante el desarrollo. Su capacidad para integrar información de movimiento local, resolver la ambigüedad del problema de la apertura y diferenciar entre el movimiento de componentes y patrones la convierte en un componente indispensable de nuestra experiencia visual dinámica.
Si quieres conocer otros artículos parecidos a El Área MT y el Misterio del Movimiento Visual puedes visitar la categoría Neurociencia.