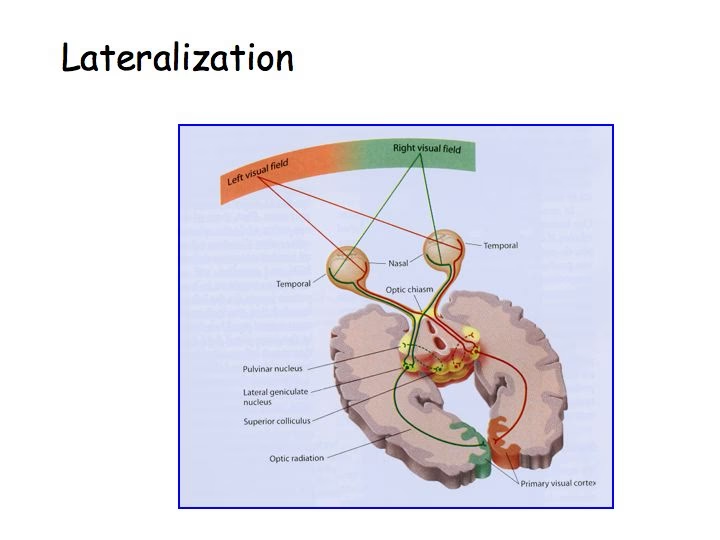
El sistema visual es una maravilla de la ingeniería biológica, una red intrincada que procesa la luz capturada por nuestros ojos para construir la rica experiencia visual que conocemos. En el corazón de esta red, actuando como una estación de retransmisión crucial y mucho más, se encuentra el núcleo geniculado lateral (NGL).
 of the Thalamus")
El NGL pertenece a la categoría de núcleos de proyección sensorial del tálamo y desempeña un papel esencial en el procesamiento visual normal. Su ubicación es estratégica: se encuentra en la región posteroventral de los núcleos talámicos, inmediatamente adyacente al pulvinar y posterior al punto coroideo inferior del plexo coroideo. El nombre 'geniculado' proviene del latín 'geniculum', que significa 'articulación' o 'doblez', refiriéndose a la forma curva de sus láminas, mientras que 'lateral' lo distingue del núcleo geniculado medial.
Aunque históricamente fue considerado poco más que un simple repetidor de señales, investigaciones más recientes han revelado una función mucho más compleja y dinámica para el NGL en el procesamiento visual.
- Estructura Intrincada: Láminas y Tipos Celulares
- Conexiones Vitales: Entradas y Salidas
- Más Allá del Simple Relé: Funciones Complejas
- Campos Receptivos: La Base del Procesamiento
- El NGL en la Vía Visual Retino-Genículo-Cortical
- NGL vs. V1: Diferencias Clave
- El NGL en Roedores: Subdivisiones Funcionales
- Preguntas Frecuentes sobre el NGL
- Conclusión
Estructura Intrincada: Láminas y Tipos Celulares
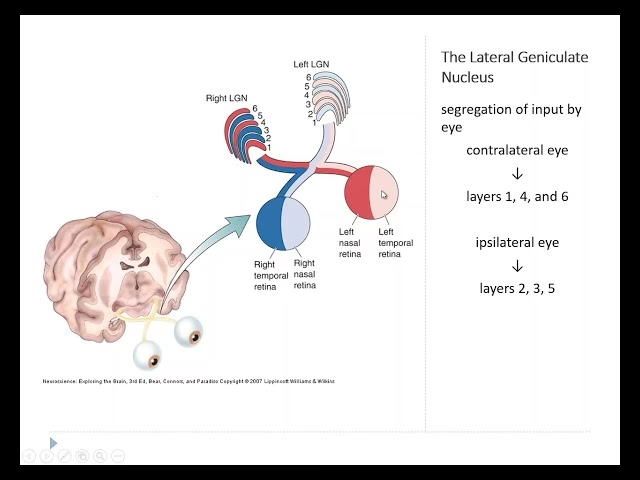
La estructura del núcleo geniculado lateral se organiza principalmente en torno a tres tipos celulares distintos: magnocelulares (M), parvocelulares (P) y koniocelulares (K). En humanos y muchos otros primates, las células P y M se disponen en seis capas diferentes (cuatro capas dorsales P y dos capas ventrales M). La información visual llega a estas capas de manera segregada según el ojo de origen:
- Las señales de los ganglios retinianos del ojo ipsilateral (el del mismo lado del cerebro) hacen sinapsis en las capas 2, 3 y 5.
- Las señales del ojo contralateral (el del lado opuesto) hacen sinapsis en las capas 1, 4 y 6.
Esta organización monocular por capa es una característica distintiva del NGL y es fundamental para el posterior procesamiento binocular en la corteza visual.
Las células M del NGL reciben información de las células ganglionares retinianas de tipo Y, que tienen campos grandes y son sensibles al movimiento. Por otro lado, las células P reciben información de las células ganglionares retinianas de tipo X, que tienen campos pequeños y son sensibles al color. Las células koniocelulares (K) se proyectan en regiones ventrales a cada una de las láminas P y M, constituyendo capas intercaladas.
El volumen promedio de cada NGL en un adulto humano es de aproximadamente 118 mm³, aunque existe variabilidad individual. En un estudio, las capas magnocelulares representaron aproximadamente 28 mm³ en total, mientras que las parvocelulares constituyeron alrededor de 90 mm³.
La Matriz Extracelular del NGL
La matriz extracelular del NGL se caracteriza por una menor presencia de las típicas redes perineuronales basadas en agrecano. En cambio, muestra una alta densidad de 'capas axonales', una estructura relacionada con una matriz más localizada en las dendritas. Esto sugiere una estrategia organizativa diferente, posiblemente especializada para un procesamiento sensorial rápido.
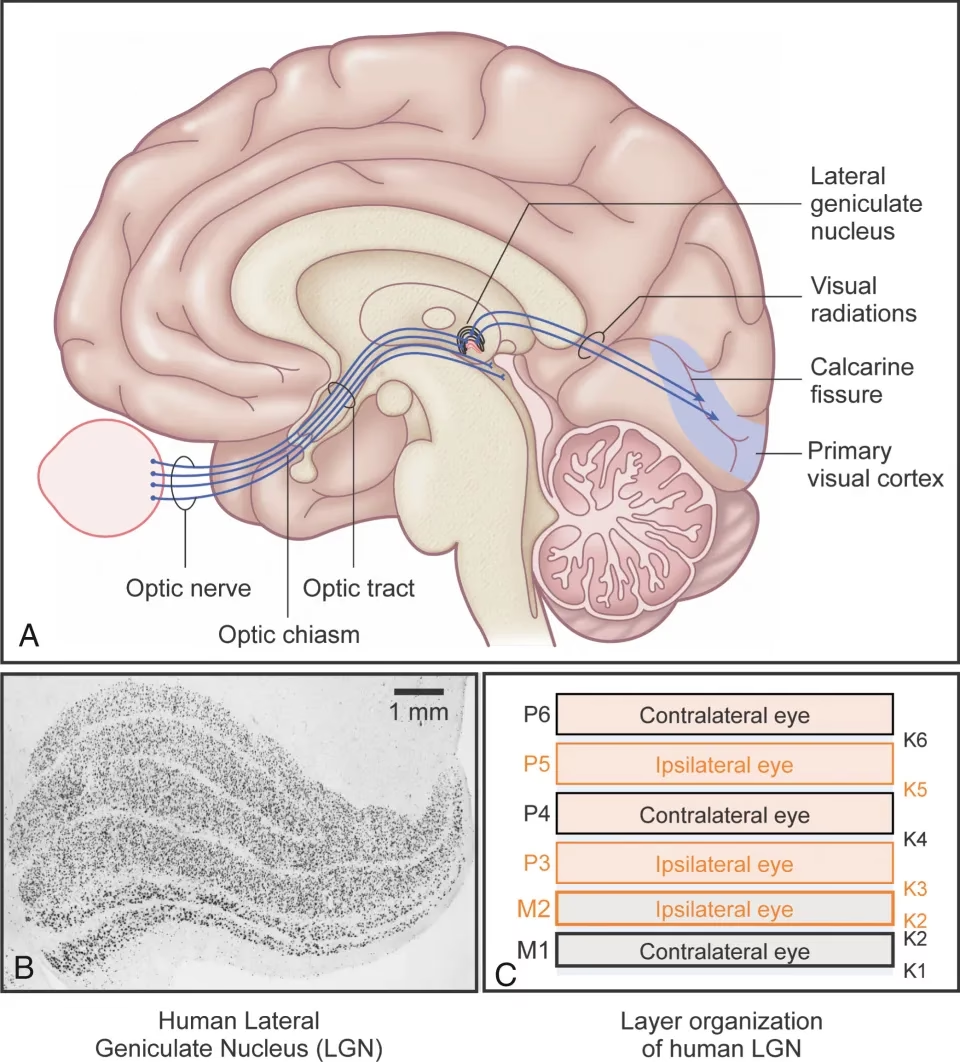
El Folículo Intergeniculado (FIG)
El NGL también contiene una sección distinta entre sus regiones dorsal y ventral conocida como el folículo intergeniculado (FIG). El FIG se proyecta a los núcleos supraquiasmáticos del hipotálamo a través del tracto geniculohipotalámico y a la glándula pineal a través del tracto geniculopineal. Esta conexión implica al NGL en la modulación de los ritmos circadianos.
Conexiones Vitales: Entradas y Salidas
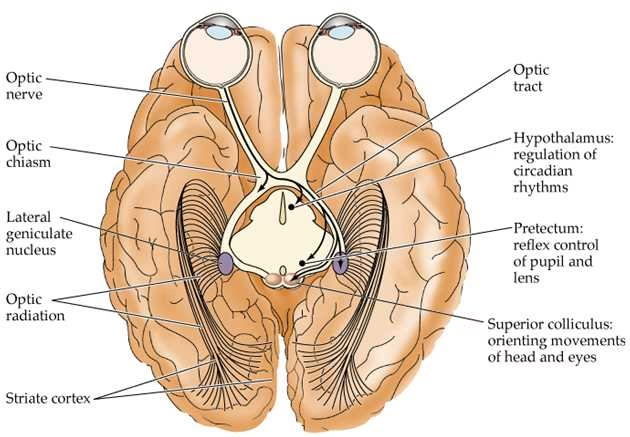
Aunque el NGL recibe una entrada considerable de los ganglios retinianos, la innervación que recibe de regiones de orden superior es mucho mayor. De hecho, hasta el 95% de la entrada al NGL proviene de la corteza visual, el colículo superior, el pretectum, los núcleos reticulares talámicos y las interneuronas locales del NGL. También recibe actividad moduladora en diversos grados de núcleos del tronco encefálico no involucrados directamente en la percepción visual, como los núcleos del rafe (serotoninérgicos), los núcleos pedunculopontino y tegmental laterodorsal (colinérgicos), y el locus coeruleus (noradrenérgicos). Estas entradas no retinianas pueden ser excitadoras, inhibidoras o moduladoras.
La información que sale del NGL viaja a través de las radiaciones ópticas (asa de Meyer, haz central y asa de Baum), que se proyectan a través de la cápsula interna hacia la corteza visual primaria (V1). Principalmente, hacen sinapsis con neuronas estrelladas espinosas en las capas 4C-alfa (recibiendo entrada magnocelular) y 4C-beta (recibiendo entrada parvocelular) de V1.
El análisis de la actividad dependiente del NGL mediante fMRI en la corteza extraestriada no-V1 sugiere que el NGL también se proyecta a regiones más downstream en la vía visual (por ejemplo, V2-V5). Además, la capa 6 de la corteza visual envía retroalimentación moduladora al NGL, al igual que el núcleo reticular talámico.
Más Allá del Simple Relé: Funciones Complejas
Como se mencionó, la visión histórica del NGL como un simple 'repetidor de señales' propuesta por Glees y LeGros Clark ha sido superada. Investigaciones posteriores han sugerido funciones más complejas, incluyendo:
- Modulación Atencional: El NGL probablemente ayuda al sistema visual a enfocar su atención en la información más importante, posiblemente influenciado por entradas moduladoras del tronco encefálico y la corteza.
- Decorrelación Temporal: Mientras que la retina realiza decorrelación espacial a través de la inhibición centro-periferia, el NGL parece lograr decorrelación temporal. Esta decorrelación espacio-temporal contribuye a una codificación mucho más eficiente de la información visual.
- Facilitación o Supresión Binocular: Mediante la modulación de ganancia monocular, el NGL puede facilitar o suprimir la información de un ojo en relación con el otro, contribuyendo al procesamiento binocular.
- Refinamiento de Campos Receptivos: Aunque las células del NGL heredan los campos receptivos centro-periferia de las células ganglionares retinianas, el NGL es una estación que puede refinar estos campos receptivos. Modelos funcionales axiomáticos describen las células del NGL en términos de Laplacianos de Gaussianas en el dominio espacial combinados con derivadas temporales, lo que concuerda con mediciones biológicas.
- Procesamiento del Color: El NGL es fundamental en los primeros pasos del procesamiento del color, donde se crean canales oponentes que comparan señales entre los diferentes tipos de fotorreceptores. Las células P contribuyen a las señales oponentes rojo-verde, las células K a las señales oponentes azul-amarillo, y las células M principalmente a la luminancia.
- Posible Sensibilidad a la Orientación: Algunas investigaciones sugieren que una subpoblación de células K en el NGL puede demostrar sensibilidad selectiva a la orientación del estímulo, similar a las células de V1.
Campos Receptivos: La Base del Procesamiento
Un concepto fundamental en las vías sensoriales tempranas es el del campo receptivo local de una neurona. En el sistema visual, el campo receptivo clásico de una neurona es la región del espacio visual que influye en la tasa de disparo de la neurona, junto con una descripción de las propiedades estructurales de ese espacio que excitan o inhiben la célula.
Las células ganglionares retinianas, que son la fuente principal de entrada visual al NGL, tienen campos receptivos con una estructura de centro-periferia (center-surround). Estos campos son circularmente simétricos y tienen una región excitatoria y una región inhibitoria. Existen dos variedades: centro ON/periferia OFF (se excitan por luz en el centro e inhiben por luz en la periferia) y centro OFF/periferia ON (se inhiben por luz en el centro y excitan por luz en la periferia). El NGL hereda esta estructura de campo receptivo de la retina.

En contraste, las células de la corteza visual primaria (V1) desarrollan campos receptivos más complejos, como los de las células simples y complejas, que son sensibles a la orientación y/o dirección del movimiento de líneas o bordes.
El NGL en la Vía Visual Retino-Genículo-Cortical
La vía visual principal desde el ojo hasta la corteza sigue el camino retina -> nervio óptico -> quiasma óptico -> tracto óptico -> NGL -> radiación óptica -> corteza visual primaria (V1).
En el quiasma óptico, las fibras de las mitades nasales (interiores) de cada retina se cruzan hacia el lado opuesto del cerebro (decusación), mientras que las fibras de las mitades temporales (exteriores) permanecen en el mismo lado. Esto significa que el NGL derecho recibe información del hemicampo visual izquierdo, y el NGL izquierdo recibe información del hemicampo visual derecho. Esta organización lateralizada se mantiene a lo largo de la vía hasta V1.
NGL vs. V1: Diferencias Clave
El NGL y la corteza visual primaria (V1) son estaciones secuenciales en la vía visual, pero tienen roles y propiedades distintivas:
| Característica | Núcleo Geniculado Lateral (NGL) | Corteza Visual Primaria (V1) |
|---|---|---|
| Ubicación | Tálamo | Corteza cerebral (Lóbulo occipital) |
| Función Principal | Retransmisión, modulación, decorrelación temporal, segregación monocular/por tipo celular, inicio procesamiento color. | Procesamiento inicial complejo, detección de características (orientación, dirección), integración binocular. |
| Campos Receptivos | Centro-periferia (heredados de la retina). | Más complejos: Células simples (sensibles a orientación con subregiones ON/OFF), Células complejas (sensibles a orientación, sin subregiones fijas, invariancia posicional limitada). |
| Organización Monocular/Binocular | Capas segregadas para cada ojo (monocular). | Neuronas binoculares (integran información de ambos ojos), organización en columnas de dominancia ocular. |
| Organización Espacial | Mapa retinotópico preservado en cada capa. | Mapa retinotópico preservado pero distorsionado (magnificación cortical de la fóvea), organización en columnas de orientación y dominancia ocular. |
| Entradas Principales | Retina (directa), Corteza visual (moduladora), Tronco encefálico (moduladora), Tálamo reticular. | NGL (entrada principal), Otras áreas corticales, Tálamo. |
| Salidas Principales | Corteza visual primaria (V1), Corteza extraestriada (V2-V5), Hipotálamo/Pineal (vía FIG). | Otras áreas visuales (V2, V3, etc.), NGL (retroalimentación). |
Mientras que el NGL mantiene la estructura básica de los campos receptivos retinianos, V1 transforma esta información para detectar características más elaboradas como la orientación de las líneas y la dirección del movimiento. La organización en columnas funcionales (dominancia ocular, orientación) es una característica prominente de V1 que no se observa de la misma manera en el NGL.
El NGL en Roedores: Subdivisiones Funcionales
En roedores, el NGL presenta subdivisiones más claras que en primates: el núcleo geniculado lateral dorsal (dorsal LGN o dLGN), el núcleo geniculado lateral ventral (ventral LGN o vLGN) y el folículo intergeniculado (IGL), ubicado entre ellos. Estas son estructuras subcorticales distintas con funciones diferenciadas.
- dLGN: Es la división principal y análoga al NGL en primates en cuanto a su rol visual. Recibe la mayoría de su entrada de la retina, está laminado y muestra organización retinotópica.
- vLGN: Es relativamente grande en varias especies, incluyendo roedores. Se divide en partes externa e interna y parece estar estrechamente relacionado con el IGL.
- IGL: Es un área pequeña dorsal al vLGN. El vLGN y el IGL comparten neuroquímicos y reciben entradas de la retina, locus coeruleus y rafe, entre otros. Han mostrado respuestas sensibles al espectro y al movimiento. Parecen desempeñar un papel importante en la mediación de las fases de los ritmos circadianos que no dependen de la luz, así como en los cambios de fase dependientes de la luz.
Preguntas Frecuentes sobre el NGL
¿Qué significa NGL en neurociencia?
NGL significa Núcleo Geniculado Lateral. Es una estructura clave ubicada en el tálamo del cerebro, parte fundamental de la vía visual.
¿Se encuentra el NGL en la retina?
No, el NGL no se encuentra en la retina. La retina es la capa sensible a la luz en la parte posterior del ojo. El NGL es una estructura cerebral, ubicada en el tálamo, que recibe información de la retina a través del nervio óptico.
¿Cuál es la función principal del Núcleo Geniculado Lateral?
Aunque inicialmente se pensó que era solo un relé, la función principal del NGL es recibir, procesar y retransmitir información visual desde la retina a la corteza visual. Participa en la segregación de la información por ojo y tipo celular, la decorrelación temporal, la modulación atencional, el procesamiento inicial del color y, a través de su folículo intergeniculado, en la modulación de los ritmos circadianos.
¿Cuáles son los tipos de células principales en el NGL?
Los tipos de células principales en el NGL son las células magnocelulares (M), parvocelulares (P) y koniocelulares (K), cada una con diferentes características y roles en el procesamiento de la información visual.
Conclusión
El núcleo geniculado lateral es una estructura fascinante y fundamental en el sistema visual. Lejos de ser un simple punto de paso, actúa como un centro de procesamiento dinámico que recibe influencias de múltiples áreas cerebrales, modulando la información visual antes de enviarla a la corteza. Su compleja organización en capas y tipos celulares, junto con sus diversas conexiones, subraya su papel esencial no solo en la percepción visual detallada, sino también en aspectos como la atención y la regulación de nuestros relojes biológicos. Comprender el NGL es adentrarse en la sofisticada forma en que nuestro cerebro construye nuestra realidad visual.
Si quieres conocer otros artículos parecidos a El Núcleo Geniculado Lateral y la Visión puedes visitar la categoría Neurociencia.