
Durante mucho tiempo, se creyó que el comportamiento de las neuronas, especialmente cómo integraban sus múltiples entradas sinápticas para generar patrones de disparo específicos, podía explicarse casi completamente a través de modelos de cable pasivos. Estos modelos veían a la neurona como un simple conducto eléctrico, donde las señales se propagaban y sumaban linealmente. Sin embargo, las últimas décadas de investigación en neurociencia han revelado una imagen mucho más compleja y fascinante: las neuronas y sus intrincados árboles dendríticos no actúan como cables simples. En cambio, exhiben una rica variedad de propiedades de membrana no lineales, que son fundamentales para comprender la relación entrada/salida de la célula neuronal.
Estas no linealidades de la membrana se deben a diversas conductancias de membrana. Una conductancia (el inverso de la resistencia) es, en esencia, la facilidad con la que las cargas eléctricas (iones) pueden fluir a través de la membrana celular. Esta capacidad de flujo está controlada por proteínas especializadas incrustadas en la membrana: los canales iónicos. Estos canales pueden abrirse o cerrarse, alterando drásticamente el flujo de iones dentro o fuera de la célula. Los cambios en la conductancia no solo afectan el potencial de membrana, sino también la resistencia de la membrana y, por lo tanto, las propiedades pasivas de cable de la célula, como la longitud electrotónica.
Consideremos, por ejemplo, una interneurona. Un modelo de cable basado en una resistencia de membrana fija predice una longitud electrotónica constante. Si esta resistencia puede variar debido a la activación o inactivación de diferentes conductancias, entonces la longitud electrotónica también puede cambiar. En un estado donde ciertas conductancias (como algunas de K+) están activas, la membrana es más permeable (mayor conductancia, menor resistencia), haciendo que la célula sea electrotónicamente más extensa. Esto podría aislar funcionalmente las entradas sinápticas distales del soma. Si estas conductancias se inactivan, la resistencia aumenta, la célula se vuelve electrotónicamente más compacta y el aislamiento se reduce.
- Fundamentos: Conductancia, Resistencia y Canales Iónicos
- Conductancias Dependientes de Voltaje: El Potencial de Acción Clásico
- Más Allá del Potencial de Acción: Diversidad de Conductancias
- La Conductancia de Calcio de Bajo Umbral (IT)
- Conductancias Asociadas y Ritmicidad
- Otras Conductancias Notables
- Impacto en la Integración Sináptica y los Patrones de Disparo
- Tabla Comparativa de Conductancias Clave
- Preguntas Frecuentes
Fundamentos: Conductancia, Resistencia y Canales Iónicos
La base de la excitabilidad neuronal reside en la capacidad de la membrana celular para controlar el flujo de iones. Las diferencias de concentración de iones a través de la membrana, mantenidas activamente por bombas iónicas (como la bomba de sodio-potasio), crean gradientes electroquímicos. Sin embargo, la membrana no es perfectamente impermeable. Existe una "fuga" constante de iones a través de canales de fuga, que generalmente no dependen del voltaje de la membrana. Esta fuga, predominantemente de iones K+, combinada con la acción de las bombas, establece el potencial de membrana en reposo, típicamente entre -65 y -75 mV en muchas neuronas.
La conductancia de la membrana a un ion particular está directamente relacionada con el número de canales iónicos abiertos permeables a ese ion. Una mayor conductancia significa que más iones pueden cruzar la membrana por unidad de tiempo para un dado gradiente electroquímico. La resistencia de la membrana es simplemente el inverso de la conductancia total a todos los iones permeables. Los cambios en la conductancia, por ejemplo, debido a la apertura de canales iónicos en respuesta a una entrada sináptica (canales activados por ligando) o a un cambio en el voltaje de la membrana (canales activados por voltaje), son los eventos fundamentales que subyacen a la señalización neuronal.
Conductancias Dependientes de Voltaje: El Potencial de Acción Clásico
El ejemplo más conocido de conductancias dependientes de voltaje es el que subyace al potencial de acción, descrito clásicamente por Hodgkin y Huxley en el axón gigante de calamar. El potencial de acción es un evento de todo o nada que permite la propagación rápida de señales a largas distancias a lo largo del axón.
En el potencial de acción típico, intervienen principalmente dos tipos de conductancias dependientes de voltaje: la conductancia al Na+ y la conductancia al K+.
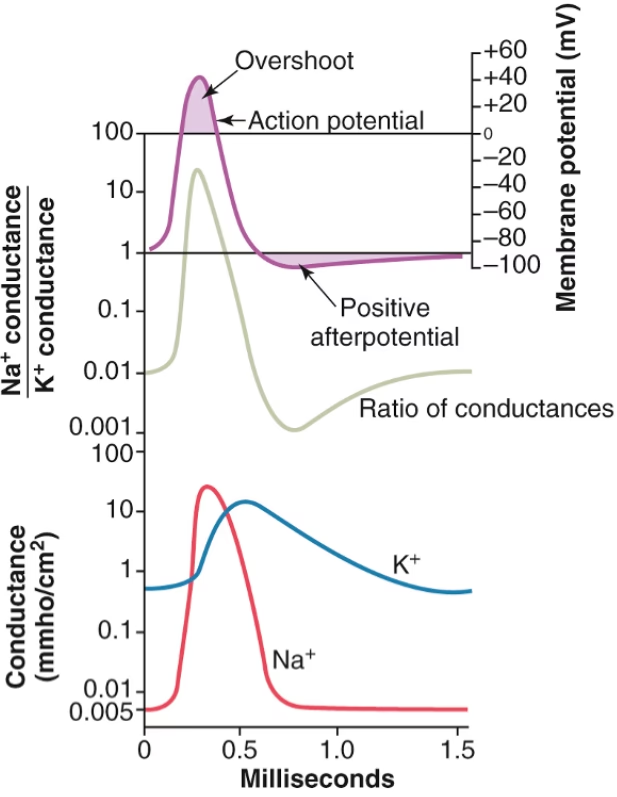
- Conductancia al Na+: Los canales de Na+ dependientes de voltaje están cerrados en reposo. Cuando el potencial de membrana alcanza un cierto umbral de despolarización (alrededor de -50 mV desde un potencial de reposo de -70 mV), estos canales se abren rápida y masivamente. Esto aumenta bruscamente la conductancia al Na+, permitiendo que los iones Na+ fluyan hacia la célula, impulsados por su gradiente electroquímico. Esta afluencia rápida de carga positiva causa la fase ascendente (despolarización) del potencial de acción. Sin embargo, esta apertura es transitoria. Poco después de abrirse, los canales de Na+ entran en un estado inactivado, donde permanecen cerrados independientemente del voltaje. La inactivación requiere que la célula se repolarice a niveles más negativos para ser eliminada (desinactivación).
- Conductancia al K+: Los canales de K+ dependientes de voltaje también se abren en respuesta a la despolarización, pero con una cinética más lenta que los canales de Na+. Su apertura aumenta la conductancia al K+, permitiendo que los iones K+ fluyan fuera de la célula (impulsados por su gradiente). Este eflujo de carga positiva causa la fase descendente (repolarización) del potencial de acción, ayudando a devolver el potencial de membrana a su nivel de reposo. A diferencia de los canales de Na+, estos canales de K+ clásicos no muestran una inactivación significativa dependiente de la despolarización; simplemente se cierran a medida que el potencial de membrana se hiperpolariza (desactivación).
La interacción temporal y dependiente del voltaje de estas dos conductancias genera la forma característica del potencial de acción, incluyendo un período de hiperpolarización posterior (poshiperpolarización). La inactivación de los canales de Na+ y la activación lenta de los canales de K+ son responsables del período refractario, que limita la frecuencia máxima de disparo neuronal y asegura la propagación unidireccional del potencial de acción a lo largo del axón.
La propagación del potencial de acción, conocida como conducción nerviosa, es un resultado directo de estas conductancias dependientes de voltaje. La despolarización local causada por la entrada de Na+ activa canales de Na+ vecinos, propagando la onda de despolarización a lo largo del axón. En axones mielinizados, los canales de Na+ y K+ se concentran en los Nodos de Ranvier, lo que permite una conducción saltatoria mucho más rápida, donde el potencial de acción "salta" de nodo en nodo.
Más Allá del Potencial de Acción: Diversidad de Conductancias
Si bien las conductancias de Na+ y K+ subyacentes al potencial de acción clásico son fundamentales, las neuronas, especialmente en el sistema nervioso central de mamíferos, poseen una asombrosa variedad de otras conductancias de membrana. Muchas de ellas también son dependientes de voltaje, pero difieren en sus umbrales de activación, cinéticas, selectividad iónica y ubicación subcelular (soma, dendritas, axón, terminales sinápticas). Además, algunas conductancias son activadas o moduladas por la concentración intracelular de iones (como Ca2+) o por la acción de neurotransmisores a través de receptores metabotrópicos.
Esta diversidad de conductancias es lo que permite a las neuronas exhibir patrones de disparo complejos y dinámicos, y lo que hace que la integración sináptica sea un proceso altamente no lineal y sofisticado, mucho más allá de lo que predice un simple modelo de cable pasivo.
La Conductancia de Calcio de Bajo Umbral (IT)
Una de las conductancias dependientes de voltaje más importantes, particularmente en las neuronas de relevo talámico, es la conductancia de Ca2+ de bajo umbral, mediada por los canales de Calcio tipo T ("T" por transitorio). Esta conductancia (y la corriente asociada, IT) es crucial para determinar si una neurona de relevo talámico responde a una entrada en modo tónico o en modo de ráfaga.
Similar a la conductancia de Na+ del potencial de acción, la IT tiene estados de activación, inactivación y desinactivación dependientes del voltaje. Sin embargo, su umbral de activación está a un potencial de membrana más hiperpolarizado (más negativo) que el del potencial de acción. La IT está inactivada cuando la membrana está relativamente despolarizada (más positiva que aproximadamente -60 mV). Pero se desinactiva cuando la membrana se hiperpolariza (más negativa que aproximadamente -60 mV). Una vez desinactivada, una despolarización suficiente (como un potencial postsináptico excitador - EPSP) puede activar la IT.
La activación de la IT provoca una entrada de iones Ca2+ en la célula, generando una despolarización distintiva en forma de pico, conocida como el pico de bajo umbral (LTS - Low-Threshold Spike). Este LTS es un evento regenerativo y, en neuronas no sometidas a pinzamiento de voltaje (voltage clamp), se comporta de manera similar a un evento de "todo o nada". Una vez que se alcanza el umbral para activar la IT, incluso una pequeña despolarización adicional evoca un LTS de amplitud máxima, aunque su latencia puede disminuir con estímulos más fuertes.
El LTS es significativo porque, al ser una gran despolarización (20-30 mV), es capaz de activar los canales de Na+ y K+ dependientes de voltaje que generan los potenciales de acción convencionales. Así, un único LTS puede desencadenar una breve ráfaga (burst) de potenciales de acción (generalmente entre 2 y 10).
Modos de Disparo: Tónico vs. Ráfaga
La IT es el interruptor entre el modo tónico y el modo de ráfaga:
- Modo Tónico: Ocurre cuando la membrana está relativamente despolarizada (por ejemplo, durante el estado de vigilia atenta). En este estado, la IT está inactivada y no puede ser activada por las entradas sinápticas. La neurona se comporta de manera más similar a un integrador lineal; su respuesta (frecuencia de disparo) se relaciona de forma más lineal con la fuerza y duración de la entrada despolarizante.
- Modo de Ráfaga: Ocurre cuando la membrana está relativamente hiperpolarizada (por ejemplo, durante el sueño de ondas lentas o la somnolencia). En este estado, la IT está desinactivada. Una entrada despolarizante (un EPSP) que alcance el umbral activará el LTS. El LTS, a su vez, desencadena una ráfaga de potenciales de acción. La relación entre la entrada y la respuesta de ráfaga es altamente no lineal; una entrada que apenas supera el umbral puede evocar una ráfaga completa, mientras que entradas más fuertes pueden no aumentar mucho el número de espigas dentro de la ráfaga inicial, aunque pueden acortar su latencia.
La IT también tiene una dependencia temporal notable. La inactivación después de la despolarización y la desinactivación después de la hiperpolarización son procesos relativamente lentos, a menudo requiriendo 100 ms o más. Esto impone un período refractario al LTS, limitando la frecuencia de los picos de bajo umbral a 10 Hz o menos. Este lento proceso contribuye a la naturaleza rítmica del disparo en modo de ráfaga observado en ciertas condiciones.
Conductancias Asociadas y Ritmicidad
Además de la IT, otras conductancias interactúan para dar forma a los patrones de disparo, especialmente en el modo de ráfaga y la generación de ritmos:
- Conductancias de K+ Dependientes de Voltaje y/o Ca2+: La gran despolarización del LTS y la entrada de Ca2+ asociada pueden activar varias conductancias de K+. Estas contribuyen a la repolarización rápida de la membrana después del LTS y la ráfaga de potenciales de acción, ayudando a devolver la célula a un estado hiperpolarizado desde el cual la IT puede desinactivarse nuevamente.
- Conductancia Catiónica Activada por Hiperpolarización (Ih): Esta conductancia (también conocida como "corriente de sag") se activa por la hiperpolarización de la membrana y se desactiva por la despolarización. Permite la entrada de cationes (Na+ y K+), generando una corriente despolarizante lenta. La activación de Ih es notablemente lenta (> 200 ms).
La interacción cíclica entre la IT, las conductancias de K+ asociadas y la Ih puede generar un disparo rítmico de ráfagas. Una hiperpolarización inicial desinactiva la IT. La hiperpolarización también activa lentamente la Ih, que despolariza la célula. Esta despolarización, si alcanza el umbral, activa la IT, generando un LTS y una ráfaga. El LTS y la ráfaga activan conductancias de K+ que hiperpolarizan la célula, inactivando la IT y desactivando la Ih. La hiperpolarización sostenida desinactiva nuevamente la IT y activa lentamente la Ih, reiniciando el ciclo. Este mecanismo contribuye a los ritmos talámicos observados durante el sueño.
Interesantemente, la Ih puede ser modulada por neuromoduladores como la serotonina, noradrenalina e histamina, lo que permite que el estado fisiológico general del cerebro influya en los patrones de disparo neuronal.
Otras Conductancias Notables
Existen otras conductancias importantes que contribuyen a la dinámica neuronal:
- Conductancia IA: Es una conductancia de K+ dependiente de voltaje con una cinética de inactivación rápida y un umbral de activación relativamente bajo (aunque más despolarizado que la IT). También tiene estados de activación, inactivación y desinactivación. A diferencia de la IT (que despolariza), la IA hiperpolariza. Se sugiere que la IA puede extender el rango dinámico de respuesta de la neurona durante el modo tónico, ralentizando y reduciendo la despolarización causada por los EPSPs, evitando que la célula alcance la saturación de disparo demasiado pronto.
- Conductancias de Calcio de Alto Umbral: Además de la IT, existen conductancias de Ca2+ que requieren una despolarización mucho mayor para activarse (alrededor de -20 mV). Incluyen los canales tipo L ("long-lasting", inactivación lenta) y tipo N ("neither T nor L", inactivación más rápida que L). Estos canales se localizan a menudo en dendritas y, crucialmente, en terminales sinápticas. En los terminales, la entrada de Ca2+ a través de estos canales, activada por la llegada de un potencial de acción, es el paso esencial que desencadena la liberación de neurotransmisores. En las dendritas, pueden contribuir a la integración de entradas sinápticas distales o a la "retropropagación" del potencial de acción desde el soma.
- Conductancias de K+ Dependientes de Ca2+: Distintas a las asociadas al LTS rítmico, existen otras conductancias de K+ activadas por un aumento en la concentración intracelular de Ca2+. Este aumento a menudo es el resultado de la activación de conductancias de Ca2+ de alto umbral durante un potencial de acción. Algunas son rápidas y contribuyen a la repolarización y poshiperpolarización. Otras son más lentas y se acumulan con potenciales de acción repetidos, llevando a la adaptación de la frecuencia de disparo: una reducción gradual en la frecuencia de los potenciales de acción a pesar de un estímulo constante.
Impacto en la Integración Sináptica y los Patrones de Disparo
La compleja interacción de todas estas conductancias determina cómo una neurona integra las miríadas de entradas sinápticas que recibe y cuál será su patrón de disparo de salida. Ya no podemos ver a la neurona como un simple sumador lineal.
Las entradas sinápticas, al abrir canales iónicos activados por ligando, causan cambios en la conductancia que alteran el potencial de membrana (EPSPs o IPSPs). Sin embargo, estos cambios en el potencial de membrana pueden, a su vez, activar o inactivar las conductancias dependientes de voltaje intrínsecas de la neurona. Esto crea un ciclo de retroalimentación donde las entradas sinápticas modulan las propiedades de membrana intrínsecas de la célula, que a su vez influyen en cómo la célula responde a futuras entradas.

Por ejemplo, el estado de polarización de la membrana (y, por lo tanto, el estado de inactivación/desinactivación de la IT) determina si una entrada despolarizante resultará en un disparo tónico lineal o en una ráfaga no lineal. La presencia de la IA puede modificar la respuesta tónica. Las conductancias dendríticas pueden alterar la forma en que las entradas distales llegan al soma. Las conductancias dependientes de Ca2+ pueden limitar la duración de una descarga o inducir adaptación.
Comprender estas conductancias es esencial para desentrañar la complejidad de la computación neuronal y cómo los diferentes tipos de neuronas generan sus patrones de actividad característicos que subyacen a la función cerebral, desde el procesamiento sensorial en el tálamo hasta la toma de decisiones en la corteza.
Tabla Comparativa de Conductancias Clave
| Conductancia | Ion Principal | Dependencia Principal | Función Clave |
|---|---|---|---|
| Fuga de K+ | K+ | Independiente de voltaje | Establecer potencial de reposo |
| Na+ (Potencial de Acción) | Na+ | Voltaje (alto umbral despol.) | Fase ascendente del potencial de acción |
| K+ (Potencial de Acción) | K+ | Voltaje (lento, despol.) | Fase descendente/repolarización del potencial de acción |
| Ca2+ de Bajo Umbral (IT) | Ca2+ | Voltaje (bajo umbral despol., inactivación por despol.) | Generar LTS, control de modo tónico/ráfaga |
| Ih | Na+, K+ | Hiperpolarización | "Corriente de sag", contribuye a ritmos |
| IA | K+ | Voltaje (despol., inactivación rápida) | Modula respuesta tónica, posible rango dinámico |
| Ca2+ de Alto Umbral | Ca2+ | Voltaje (alto umbral despol.) | Liberación de neurotransmisores, integración dendrítica |
| K+ Dependiente de Ca2+ | K+ | Ca2+ intracelular | Repolarización, poshiperpolarización, adaptación de frecuencia |
Preguntas Frecuentes
¿Qué diferencia hay entre la conducción nerviosa y la conductancia neuronal?
La conductancia neuronal se refiere a la capacidad de la membrana de una neurona para permitir el paso de iones a través de canales, lo que genera corrientes eléctricas y cambia el potencial de membrana. La conducción nerviosa, o propagación del potencial de acción, es el fenómeno de cómo una señal eléctrica (el potencial de acción) viaja a lo largo del axón. La conducción nerviosa es un *resultado* de los cambios coordinados en las conductancias dependientes de voltaje a lo largo de la membrana del axón.
¿Cómo influye la conductancia de calcio de bajo umbral (IT) en el disparo de las neuronas?
La IT actúa como un interruptor entre dos modos de disparo principales en neuronas como las talámicas: el modo tónico y el modo de ráfaga. Cuando la membrana está despolarizada, la IT está inactivada y la neurona dispara en modo tónico, respondiendo linealmente a las entradas. Cuando la membrana está hiperpolarizada, la IT está desinactivada. Una entrada despolarizante activa la IT, generando un pico de bajo umbral (LTS) que desencadena una ráfaga de potenciales de acción. Esto permite una respuesta no lineal y amplificada a ciertas entradas.
¿Pueden cambiar las propiedades de conductancia de una neurona?
Sí, las propiedades de conductancia de una neurona no son estáticas. Pueden ser moduladas por neurotransmisores, neuromoduladores, segundos mensajeros y el historial de actividad de la neurona. Por ejemplo, la conductancia Ih puede ser modulada por neuromoduladores. La expresión y localización de los canales iónicos también pueden regularse a largo plazo, modificando fundamentalmente la excitabilidad de la célula.
¿Por qué es importante estudiar estas conductancias más allá de las del potencial de acción clásico?
Estudiar la diversidad de conductancias es crucial porque son responsables de los complejos patrones de disparo y las propiedades computacionales de las neuronas que no pueden explicarse por el simple modelo de cable o solo por el potencial de acción. Estas conductancias intrínsecas interactúan con las entradas sinápticas para determinar cómo la información se procesa, se integra y se transmite dentro de los circuitos neuronales, lo cual es fundamental para funciones cerebrales como la percepción, la atención, el aprendizaje y el control motor.
Si quieres conocer otros artículos parecidos a Conductancia Neuronal: Clave del Disparo Celular puedes visitar la categoría Neurociencia.