La capacidad de ver, de percibir el mundo que nos rodea en toda su riqueza de formas y colores, es uno de los sentidos más complejos y fascinantes. Este prodigioso acto es posible gracias a una intrincada red de estructuras neurológicas que constituyen lo que conocemos como la vía visual. Esta vía es la autopista de información que transporta las señales luminosas captadas por los ojos hasta las áreas del cerebro encargadas de interpretarlas como imágenes coherentes.

El viaje de la luz comienza mucho antes de que se convierta en una percepción consciente. Desde el momento en que los fotones inciden en la superficie del ojo, se desencadena una cascada de eventos que transforman la energía luminosa en impulsos eléctricos. Esta transducción, el primer paso crucial, ocurre en la retina, la capa más interna del ojo, que actúa como el punto de partida de esta compleja vía sensorial.

- Anatomía Detallada de la Vía Visual
- La Retina: El Punto de Inicio
- El Nervio Óptico (II Par Craneal)
- El Quiasma Óptico y los Tractos Ópticos
- El Cuerpo Geniculado Lateral (CGL)
- Las Radiaciones Ópticas
- La Corteza Visual Primaria (Área 17 de Brodmann)
- Áreas Visuales de Asociación (Áreas 18 y 19 de Brodmann y más allá)
- Campos Oculares Frontales
- Otras Vías Visuales No Conscientes
- Preguntas Frecuentes sobre la Vía Visual
- Conclusión
Anatomía Detallada de la Vía Visual
Aunque la luz interactúa primero con la córnea, la vía visual propiamente dicha se inicia en la retina. Esta vía no es un simple cable que lleva información de A a B, sino una serie de estaciones de relevo y procesamiento, cada una con su rol específico. Las estructuras principales involucradas en este recorrido incluyen los nervios ópticos, el quiasma óptico, los tractos ópticos, el cuerpo geniculado lateral, las radiaciones ópticas y, finalmente, la corteza visual, junto con sus proyecciones y áreas de asociación.
La Retina: El Punto de Inicio
La retina es una delicada capa de tejido nervioso que recubre la superficie interna posterior del globo ocular, extendiéndose hasta la ora serrata, donde se une al cuerpo ciliar. Es aquí donde la luz, tras atravesar la córnea, el cristalino y los humores acuoso y vítreo, alcanza la capa de fotorreceptores. La retina está compuesta por diez capas distintas, cada una con funciones y tipos celulares específicos. Estas capas, desde la más externa (cerca de la coroides) hasta la más interna (cerca del humor vítreo), son:
| Capa | Descripción Breve | Células Principales |
|---|---|---|
| Epitelio Pigmentado | Establece barrera retino-sanguínea, fagocitosis, protección lumínica. | Epitelio cúbico simple pigmentado. |
| Capa de Bastones y Conos | Contiene los fotorreceptores que transducen la luz. | Bastones (baja luz), Conos (alta luz, color). |
| Membrana Limitante Externa | Lámina fenestrada que une procesos de fotorreceptores. | Procesos de células de Müller. |
| Capa Nuclear Externa | Contiene los cuerpos celulares de bastones y conos. | Núcleos de bastones y conos. |
| Capa Plexiforme Externa | Sinapsis entre fotorreceptores, células bipolares y horizontales. | Axones de fotorreceptores, dendritas de bipolares y horizontales. |
| Capa Nuclear Interna | Contiene cuerpos celulares de interneuronas retinianas. | Células bipolares, horizontales y amacrinas. |
| Capa Plexiforme Interna | Sinapsis entre células bipolares/amacrinas y células ganglionares. | Axones/dendritas de bipolares, amacrinas y ganglionares. |
| Capa de Células Ganglionares | Contiene los cuerpos celulares de las neuronas de segundo orden. | Células ganglionares. |
| Capa de Fibras Nerviosas | Axones no mielinizados de las células ganglionares que forman el nervio óptico. | Axones de células ganglionares, fibras de Müller. |
| Membrana Limitante Interna | Capa más interna que cubre la superficie vítrea. | Procesos terminales de células de Müller. |
Los bastones, más numerosos, son responsables de la visión en condiciones de baja luminosidad (visión escotópica), mientras que los conos, concentrados en la fóvea, permiten la visión de alta resolución y la percepción del color (visión fotópica). La fóvea, una pequeña depresión en el centro de la mácula (una zona amarillenta lateral al disco óptico), es la región de máxima agudeza visual debido a la alta densidad de conos y la reducción del espesor de las capas internas que permiten que la luz llegue más directamente a los fotorreceptores. El disco óptico, por otro lado, carece de fotorreceptores y es el punto ciego fisiológico.
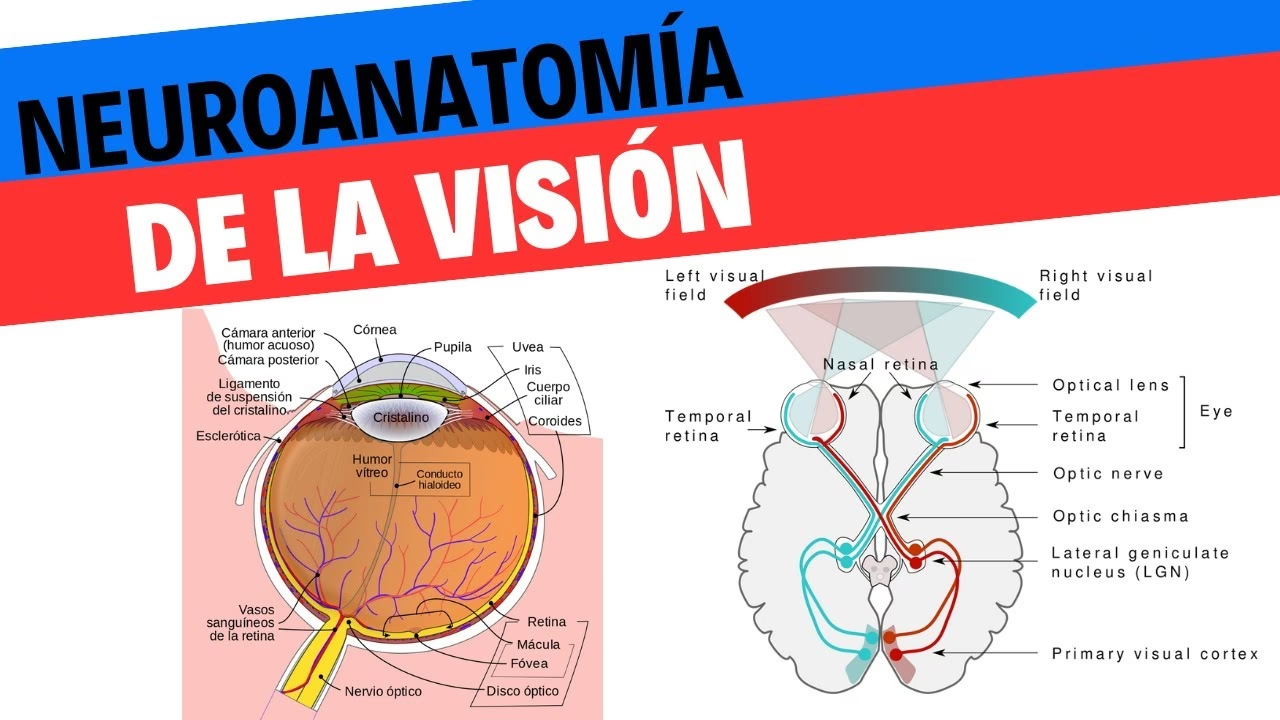
Es fundamental comprender la organización retinotópica: la retina se divide en cuadrantes (nasal superior/inferior, temporal superior/inferior). La porción nasal de una retina y la porción temporal de la retina contralateral reciben información del mismo hemicampo visual. Específicamente, el hemicampo visual izquierdo se proyecta a la porción nasal de la retina izquierda y a la porción temporal de la retina derecha. El hemicampo visual derecho se proyecta a la porción nasal de la retina derecha y a la porción temporal de la retina izquierda.
El Nervio Óptico (II Par Craneal)
Los axones de aproximadamente un millón de células ganglionares convergen en el disco óptico para formar el nervio óptico. Estos axones, a diferencia de los fotorreceptores, están mielinizados, aunque por oligodendrocitos (células gliales del sistema nervioso central), lo que subraya que el nervio óptico es funcionalmente una extensión del SNC. El nervio óptico sale del ojo, atraviesa la órbita a través de la fisura orbitaria superior y se dirige hacia el encéfalo. A lo largo de su recorrido, está rodeado por las meninges (duramadre y aracnoides), que están en continuidad con las del cerebro. Se subdivide en porciones: cabeza del nervio óptico (dentro del ojo), porción intraorbitaria, porción intracanalicular (en el canal óptico) y porción intracraneal.
El Quiasma Óptico y los Tractos Ópticos
Los dos nervios ópticos se encuentran por encima del diafragma sellar para formar el quiasma óptico. Esta estructura es un punto crucial no solo de unión, sino de decusación (cruce) parcial. Aquí, las fibras nerviosas procedentes de las porciones nasales de ambas retinas se cruzan hacia el lado contralateral, mientras que las fibras procedentes de las porciones temporales de ambas retinas continúan su trayecto sin cruzarse (fibras ipsilaterales). Este cruce es esencial para la visión binocular.
Como resultado de esta decusación, la información del hemicampo visual izquierdo (captada por la retina nasal izquierda y la temporal derecha) viaja en el tracto óptico derecho. De manera recíproca, la información del hemicampo visual derecho (captada por la retina nasal derecha y la temporal izquierda) viaja en el tracto óptico izquierdo. Cada tracto óptico, por lo tanto, transporta la información visual de la mitad contralateral del campo visual.
Los tractos ópticos se dirigen caudalmente, rodeando los pedúnculos cerebrales del mesencéfalo rostral. Cada tracto se divide en dos raíces:
- La raíz lateral (mayor): Contiene la gran mayoría de las fibras (aproximadamente el 90%) y se dirige al cuerpo geniculado lateral del tálamo, siendo fundamental para la sensación visual consciente.
- La raíz medial (menor): Contiene aproximadamente el 10% de las fibras y se conecta con el área pretectal y el colículo superior a través del brazo superior. Estas fibras no están involucradas en la visión consciente, sino en reflejos visuales como el reflejo pupilar a la luz, el reflejo de orientación visual (movimientos automáticos de los ojos y la cabeza hacia un estímulo) y la regulación de los ritmos circadianos a través del tracto retinohipotalámico que llega al núcleo supraquiasmático del hipotálamo.
El Cuerpo Geniculado Lateral (CGL)
El cuerpo geniculado lateral es una estructura ovoide situada en la parte posterior e inferior del tálamo. Actúa como la principal estación de relevo para la información visual que se dirige a la corteza cerebral. El CGL humano está organizado en seis capas celulares, numeradas del 1 al 6 desde ventral a dorsal. La información de cada ojo se mantiene separada en capas alternas:
- Las fibras cruzadas del tracto óptico contralateral terminan en las capas 1, 4 y 6.
- Las fibras no cruzadas del tracto óptico ipsilateral terminan en las capas 2, 3 y 5.
Aunque el CGL recibe información de ambos ojos, las neuronas de cada capa responden predominantemente a un solo ojo. La organización retinotópica se mantiene en el CGL, con representaciones específicas de los cuadrantes retinianos y una representación desproporcionadamente grande de la mácula en su porción central. Además de recibir información del tracto óptico, el CGL también recibe aferencias de la corteza visual (áreas 17, 18, 19), centros oculomotores y la formación reticular, lo que sugiere un papel en la modulación de la información visual.
Las células en las capas 1 y 2 son más grandes y se conocen como células magnocelulares (vías M), especializadas en el procesamiento de información sobre movimiento y profundidad. Las células en las capas 3 a 6 son más pequeñas y se conocen como células parvocelulares (vías P), que procesan detalles finos, forma y color. Esta división en vías M y P se mantiene a lo largo de las radiaciones ópticas y en la corteza visual primaria, representando dos flujos paralelos de procesamiento de la información visual.
Las Radiaciones Ópticas
Los axones de las neuronas del cuerpo geniculado lateral forman un extenso haz de fibras conocido como las radiaciones ópticas o tracto geniculocalcarino. Estas fibras constituyen el último relevo antes de llegar a la corteza cerebral y transmiten la información visual ya procesada por el CGL.
Las radiaciones ópticas se extienden desde el CGL, pasan por la porción retrolenticular de la cápsula interna y luego se abren en forma de abanico a medida que se dirigen hacia la corteza occipital. Las fibras que transportan información de los cuadrantes inferiores de la retina (que representan el campo visual superior) realizan un extenso bucle hacia adelante y hacia abajo en el lóbulo temporal, conocido como el bucle de Meyer, antes de girar hacia atrás. Las fibras que transportan información de los cuadrantes superiores de la retina (que representan el campo visual inferior) toman un camino más directo hacia la corteza occipital.
La organización retinotópica se mantiene fielmente en las radiaciones ópticas, de modo que las lesiones en diferentes partes de este tracto provocan déficits visuales predecibles en campos visuales específicos. Debido a su extenso recorrido y a su paso cerca de estructuras vulnerables, las radiaciones ópticas son susceptibles de sufrir daños por accidentes cerebrovasculares o tumores.
La Corteza Visual Primaria (Área 17 de Brodmann)
El destino final de las radiaciones ópticas es la corteza visual primaria, también conocida como corteza estriada (debido a una banda blanquecina visible, la estría de Gennari) o área 17 de Brodmann. Se localiza principalmente en la cara medial del lóbulo occipital, en las profundidades del surco calcarino, extendiéndose hasta el polo occipital. La corteza por encima del surco calcarino es el giro Cúneo, y la de abajo es el giro Lingual.
La corteza visual primaria es la primera estación cortical donde la información visual de ambos ojos se integra a nivel neuronal. Mantiene una organización retinotópica precisa: los cuadrantes retinianos superiores (campo visual inferior) se proyectan al labio superior del surco calcarino, y los cuadrantes retinianos inferiores (campo visual superior) al labio inferior. La región macular, de alta agudeza visual, tiene una representación desproporcionadamente grande en la porción posterior del área 17.
Esta corteza está organizada en seis capas principales, con la estría de Gennari en la capa IV, que recibe la mayoría de las aferencias del CGL. Las neuronas del área 17 responden a características visuales básicas como la orientación de las líneas, los bordes y el movimiento simple. La información se organiza en columnas de dominancia ocular, donde las neuronas responden preferentemente a la entrada de un ojo u otro, y columnas de orientación, que responden a líneas con ángulos específicos. Desde el área 17, la información se proyecta a áreas visuales secundarias para un procesamiento más complejo.
Áreas Visuales de Asociación (Áreas 18 y 19 de Brodmann y más allá)
Las áreas visuales de asociación, como las áreas 18 y 19 de Brodmann, rodean la corteza visual primaria y extienden el procesamiento visual a niveles superiores. Reciben aferencias del área 17, del tálamo y de otras regiones corticales. Estas áreas no tienen la característica estría de Gennari y tienen una estructura celular ligeramente diferente.
Desde las áreas 18 y 19 (y otras áreas visuales superiores), la información se procesa a través de dos grandes vías o corrientes principales:
- La vía dorsal (también llamada 'vía del dónde'): Se proyecta hacia la corteza parietal. Está involucrada en el procesamiento de la información espacial, como la localización de objetos, el movimiento, la profundidad (estereopsis) y la coordinación sensoriomotora para guiar acciones basadas en la visión.
- La vía ventral (también llamada 'vía del qué'): Se proyecta hacia la corteza inferotemporal. Es crucial para el reconocimiento de objetos, formas, colores y caras.
Estas vías no operan de forma totalmente independiente, sino que interactúan para construir una percepción visual completa y significativa del entorno. La información entre los hemisferios cerebrales se integra a través de fibras comisurales que cruzan la línea media en el esplenio del cuerpo calloso.
Campos Oculares Frontales
Aunque no forman parte de la vía sensorial ascendente, los campos oculares frontales (áreas 6, 8 y 9 de Brodmann en el lóbulo frontal) son cruciales para el control voluntario de los movimientos oculares, particularmente los movimientos sacádicos (saltos rápidos de los ojos entre puntos de fijación). Reciben información visual procesada de las áreas visuales de asociación y proyectan al colículo superior y a los núcleos de los nervios craneales que controlan los músculos extraoculares (III, IV y VI), permitiendo la coordinación de los movimientos de los ojos y la cabeza.
Otras Vías Visuales No Conscientes
Es importante recordar que no toda la información que viaja por el tracto óptico se dirige al CGL para la visión consciente. Como se mencionó, la raíz medial del tracto óptico se proyecta a otras áreas, mediando funciones visuales no conscientes:
- Área Pretectal: Involucrada en el reflejo pupilar a la luz, controlando el tamaño de la pupila en respuesta a la intensidad lumínica.
- Colículo Superior: Parte del tectum mesencefálico, involucrado en reflejos de orientación visual, control de movimientos oculares (especialmente sacádicos) y coordinación ojo-cabeza.
- Núcleo Supraquiasmático del Hipotálamo: Recibe información a través del tracto retinohipotalámico para regular los ritmos circadianos (ciclos sueño-vigilia) y posiblemente influir en el estado de ánimo en respuesta a la luz (fotoperiodo).
Estas vías más antiguas filogenéticamente aseguran que la información visual se utilice también para regular funciones vitales y respuestas reflejas rápidas, incluso antes de que seamos conscientes de lo que estamos viendo.
Preguntas Frecuentes sobre la Vía Visual
¿Dónde comienza exactamente la vía visual?
Aunque la luz entra por la córnea, la vía visual, en términos de transducción de energía luminosa a señal neural, comienza en la retina, específicamente en la capa de fotorreceptores (bastones y conos).
¿Por qué se cruzan algunas fibras nerviosas en el quiasma óptico?
El cruce de las fibras nasales de la retina en el quiasma óptico es esencial para la visión binocular. Permite que la información de cada hemicampo visual (izquierdo o derecho) se procese en el hemisferio cerebral contralateral. Por ejemplo, todo lo que ves a tu derecha (hemicampo visual derecho) es procesado por el lado izquierdo de tu cerebro.
¿Cuál es la función principal del cuerpo geniculado lateral?
El cuerpo geniculado lateral (CGL) en el tálamo actúa como la principal estación de relevo para la información visual que se dirige a la corteza cerebral. Aquí, la información de ambos ojos se organiza en capas separadas y se inicia la segregación de las vías de procesamiento de movimiento/profundidad (magnocelular) y forma/color (parvocelular).
¿Qué papel juega la corteza visual primaria?
La corteza visual primaria (área 17) es la primera área cortical en recibir información visual del CGL. Su función principal es el procesamiento de características visuales básicas como la orientación, los bordes y el movimiento simple, manteniendo una organización retinotópica precisa.
¿Existen vías visuales que no llevan a la visión consciente?
Sí, aproximadamente el 10% de las fibras del tracto óptico se dirigen al área pretectal y al colículo superior en el mesencéfalo, así como al hipotálamo. Estas vías median reflejos visuales (como el pupilar) y regulan ritmos biológicos, operando fuera de nuestra percepción consciente.
Conclusión
La vía visual es un sistema maravillosamente organizado que transforma la energía lumínica en la rica experiencia de la visión. Desde la compleja estructura de la retina, pasando por el cruce estratégico en el quiasma óptico, el relevo en el cuerpo geniculado lateral y el procesamiento jerárquico en la corteza visual y áreas asociadas, cada componente juega un papel vital. Comprender esta vía no solo es fundamental para la neurociencia, sino que también nos ayuda a apreciar la complejidad de un sentido que a menudo damos por sentado, y a entender cómo las lesiones en diferentes puntos pueden tener consecuencias específicas en nuestra capacidad de ver el mundo.
Si quieres conocer otros artículos parecidos a El Viaje de la Luz: Vías Visuales del Cerebro puedes visitar la categoría Neurociencia.