La comprensión de cómo funciona el cerebro, esa vasta y misteriosa red de miles de millones de células, comienza con una pregunta fundamental: ¿cuál es su unidad básica de operación? A lo largo de la historia de la neurociencia, esta pregunta ha generado un debate apasionante que ha moldeado nuestra visión del sistema nervioso. Lejos de ser una simple cuestión anatómica, definir la unidad funcional es clave para desentrañar los complejos procesos fisiológicos que subyacen al pensamiento, la emoción y el comportamiento.
The Neuron Doctrine holds that the neuron is the fundamental unit anatomically. With the exception of gap or electronic junctions between neurons, the Neuron Doctrine is anatomically true. However, it appears that gap or electronic junctions in mammalian central nervous system cannot be ignored or dismissed [89].
Tradicionalmente, este debate se ha polarizado en torno a dos visiones principales: la Doctrina Neuronal y la Teoría Reticular. Ambas teorías ofrecen perspectivas distintas sobre cómo se organiza y opera el tejido nervioso, y aunque una ha prevalecido en gran medida en el ámbito anatómico, la otra conserva una profunda relevancia cuando consideramos la función y la dinámica real del cerebro.
En esencia, las nociones más fundamentales de la neurofisiología se han dicotomizado en la Doctrina Neuronal y la Teoría Reticular. La Doctrina Neuronal sostiene que la neurona es la unidad fundamental desde el punto de vista anatómico. Con la excepción de las uniones tipo gap o uniones electrónicas entre neuronas, la Doctrina Neuronal es anatómicamente cierta. Sin embargo, parece que las uniones tipo gap o uniones electrónicas en el sistema nervioso central de mamíferos no pueden ser ignoradas o descartadas. La preocupación principal surge al generalizar la Doctrina Neuronal, que es una doctrina anatómica, a una doctrina fisiológica, donde la neurona individual se considera la unidad fundamental fisiológicamente.
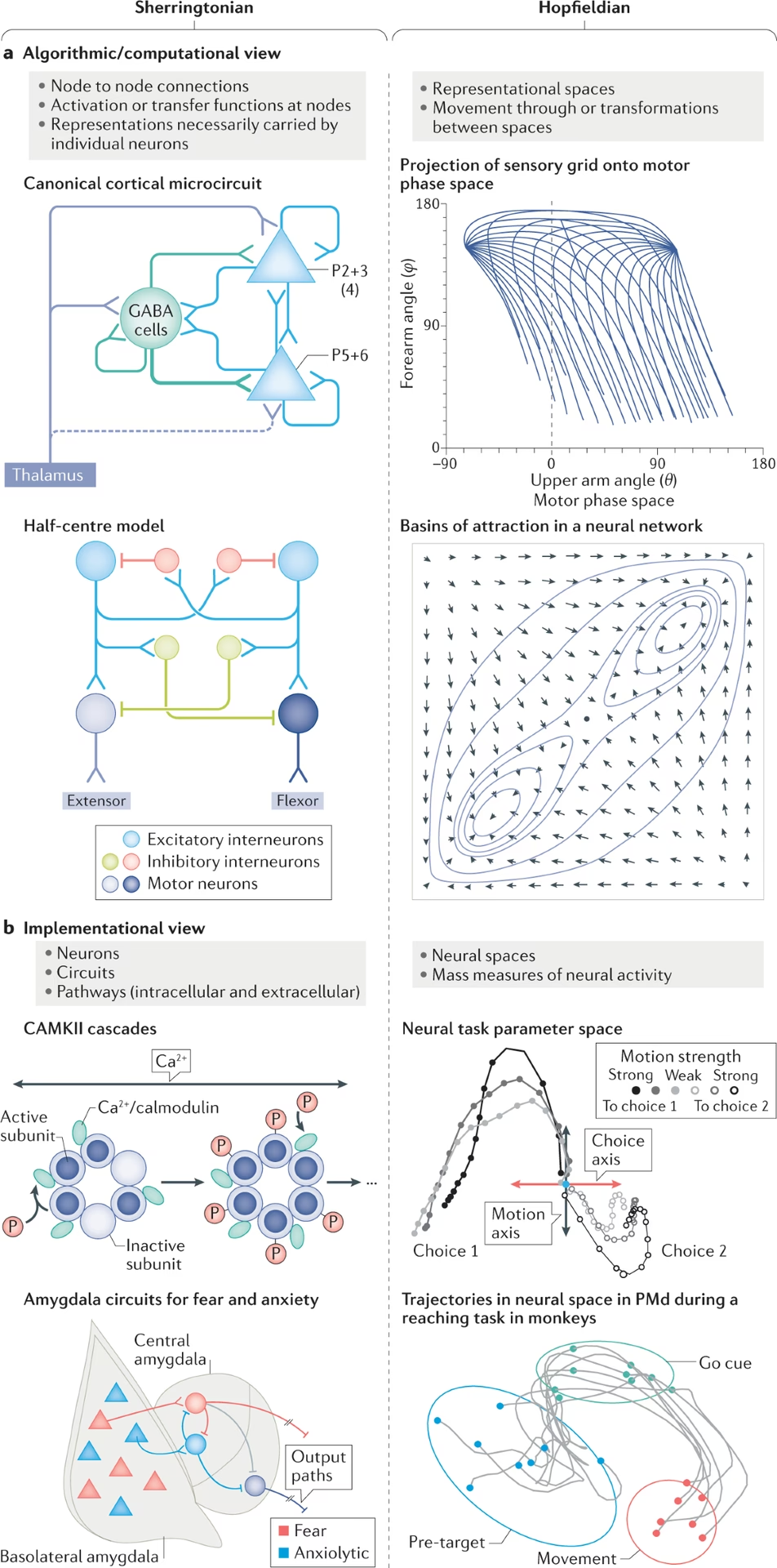
Por otro lado, los Reticularistas sostenían que la función, y por lo tanto la fisiología, no puede ser representada en la neurona individual, sino más bien en las interacciones entre un gran número de neuronas. Para el Reticularista, la unidad fundamental es la neurona Y las interacciones con las neuronas conectadas. Otros términos se han aplicado a estos conceptos, incluyendo la hipótesis de la célula cardinal, la hipótesis de acción de masa y la hipótesis conexionista.
Característica
Doctrina Neuronal
Teoría Reticular
Unidad Fundamental
La neurona individual (principalmente anatómica).
La neurona Y sus interacciones con otras neuronas (fisiológica/funcional).
Enfoque Principal
La célula nerviosa individual como entidad discreta.
La red de neuronas y las conexiones entre ellas.
Base Anatómica
Generalmente aceptada, excepto por uniones gap.
Menos énfasis en la discreción total, más en la continuidad funcional.
Base Fisiológica
Considera la neurona individual como la unidad funcional.
Considera las interacciones y la red como la base de la función.
La Naturaleza No Lineal de la Neurofisiología
Independientemente de la hipótesis que se acepte conceptualmente o predomine, cada una es dinámicamente no monótona y no lineal. Esto es un punto crucial. Una función no monótona es aquella que no es constantemente creciente ni constantemente decreciente. Una función no lineal es aquella cuya salida no es directamente proporcional a su entrada de una manera simple y predecible (no puede representarse con una línea recta o una combinación lineal simple).
La neurofisiología en su esencia es fundamentalmente no monótona y no lineal. Comprender esta naturaleza es vital para no simplificar excesivamente los procesos cerebrales.
Ejemplos de No Linealidad en Acción
Consideremos la salida del procesamiento neuronal: los potenciales de acción. La forma de un típico potencial de acción a lo largo del tiempo es claramente no monótona y no lineal. Comienza en un potencial de reposo negativo, se despolariza rápidamente hasta un pico positivo, repolariza rápidamente, a menudo hiperpolarizando brevemente por debajo del potencial de reposo, antes de volver al estado basal. Esta curva compleja no es una simple línea recta ni un cambio proporcional constante. Puede ser modelada por un conjunto de ecuaciones (como las de Hodgkin y Huxley) que son inherentemente no monótonas y no lineales.
Ahora consideremos la entrada a una neurona, como un potencial postsináptico excitatorio (PEPS) no mediado por receptores NMDA. La despolarización ascendente alcanza un pico y luego decae. Esta función también es claramente no monótona y no lineal. El PEPS puede modelarse con una ecuación como ΔVm = wt * e^(-t/tpeak), donde ΔVm es el cambio en el voltaje de la membrana en el tiempo t, tpeak es el tiempo para que el PEPS alcance su máximo, y w es una constante de proporcionalidad. Esta ecuación describe un pico seguido de una disminución exponencial, una forma característica de la no linealidad.
Luego, consideremos la propagación del PEPS a lo largo de las dendritas para alcanzar el segmento de inicio del potencial de acción. Esta propagación puede aproximarse utilizando un modelo de conducción de cable, donde el tiempo de conducción es una función de la resistencia y capacitancia de la membrana. Incluso en esta simplificación, el resultado es una función no monótona y no lineal, descrita por una ecuación diferencial parcial (como (1/ri) * (d^2V/dx^2) = Cm * (dV/dt) + V/rm, donde V es el voltaje, x la longitud, ri la resistencia axial, rm la resistencia de membrana y Cm la capacitancia de membrana). Si se consideran los diámetros cambiantes y la ramificación de las dendritas, las funciones se vuelven aún más no lineales.
Finalmente, la traducción de los PEPS iniciados en las dendritas a la generación de un potencial de acción es altamente no lineal. Una fuente de esta no linealidad es que el potencial de membrana justo antes de la llegada del PEPS afecta el umbral para generar un potencial de acción. Por ejemplo, una despolarización subumbral prolongada eleva el umbral para la generación del potencial de acción debido a una creciente inactivación de los canales iónicos de sodio (Na+). Además, los períodos refractarios después de la generación de un potencial de acción contribuyen a las no linealidades.
La Interacción Neuronal: Amplificando la Complejidad
Invocando la perspectiva Reticularista, todos estos efectos no lineales se complican aún más por el gran número de interacciones entre las neuronas conectadas. No se trata solo de la no linealidad inherente a las propiedades de una sola neurona, sino de cómo esta no linealidad se propaga y se combina a través de la vasta red. Pensemos en la dinámica donde las bolas en un juego de billar están apretadamente agrupadas antes de la ruptura; el impacto inicial (una entrada) se propaga a través de múltiples interacciones, produciendo un resultado complejo y difícil de predecir linealmente.
La Tentación de Linealizar y sus Peligros
Podría argumentarse que estas no linealidades pueden ser 'promediadas' para que se puedan realizar análisis lineales. Dicha linealización haría los análisis mucho más manejables. Uno solo necesitaría tener un tamaño de muestra suficiente de neuronas para promediar. En cierto sentido, dicho promedio sería una linealización de las funciones subyacentes no monótonas y no lineales.
Sin embargo, dicho promedio requeriría que las fases de las funciones no monótonas y no lineales fueran aleatorias e independientes. Cualquier coherencia de fase e interacción entre las neuronas en la muestra no permitiría el promedio hacia una función lineal, desde una perspectiva neurofisiológica. El problema es que no hay nada que impida a un neurocientífico utilizar métodos que linealizan el fenómeno no monótono y no lineal cuando dicha linealización no sería razonable desde una perspectiva neurofisiológica.
La tentación de linealizar funciones no monótonas y no lineales es comprensible. De hecho, la pregunta ni siquiera es considerada por muchos científicos y clínicos. Primero, para muchos científicos y clínicos es simplemente un procedimiento operativo estándar recurrir a la linealización, tanto técnica como conceptualmente. Técnicamente, las herramientas para análisis neurofisiológicos que presuponen o imponen la linealización son más numerosas y directas en sus aplicaciones. A menudo, su simplicidad es atractiva incluso cuando tergiversa la dinámica subyacente. Además, parece haber una escasez de métodos no lineales bien comprendidos y bien practicados. Más aún, la necesidad de tales métodos no lineales parece innecesaria cuando la predisposición conceptual es hacia una dinámica simple de 'empuje-tirón' (push-pull) unidimensional.
Preguntas Frecuentes
¿Cuál es la principal diferencia entre la Doctrina Neuronal y la Teoría Reticular? La Doctrina Neuronal se centra en la neurona individual como unidad anatómica. La Teoría Reticular enfatiza que la unidad funcional reside en las interacciones y la red de muchas neuronas, no solo en la neurona individual.
¿Por qué se dice que la neurofisiología es no lineal? Los procesos clave como el potencial de acción, la propagación de señales en dendritas y la traducción de entradas a salidas neuronales no siguen relaciones proporcionales simples. Sus dinámicas (cambios a lo largo del tiempo o con respecto a la entrada) implican picos, decaimientos exponenciales y umbrales que no son lineales.
¿Qué significa que sea no monótona? Significa que las funciones fisiológicas no solo aumentan o solo disminuyen; pueden subir y luego bajar, o viceversa, como se ve en la forma del potencial de acción o el PEPS.
¿Se pueden usar análisis lineales para estudiar el cerebro? Aunque es tentador por su simplicidad, los análisis lineales a menudo no son apropiados porque la neurofisiología es fundamentalmente no lineal y las interacciones neuronales introducen complejidades. Promediar datos para linealizar solo funciona si las dinámicas subyacentes son aleatorias e independientes, lo cual no suele ser el caso en las redes neuronales activas. Usar métodos lineales puede tergiversar la verdadera dinámica cerebral.
Si quieres conocer otros artículos parecidos a Debate Fundamental en Neurociencia puedes visitar la categoría Neurociencia.
Soy licenciado en Medicina y Cirugía y Doctor en Neurociencias por la Universidad Complutense de Madrid. Me formé como especialista en Neurología realizando la residencia en el Hospital 12 de Octubre bajo la dirección de Alberto Portera y Alfonso Vallejo, donde también ejercí como adjunto durante seis años y fui tutor de residentes. Durante mi formación, realicé una rotación electiva en el Memorial Sloan Kettering Cancer Center.Posteriormente, fui Jefe de Sección en el Hospital Clínico San Carlos de Madrid y actualmente soy jefe de servicio de Neurología en el Hospital Universitario Fundación Jiménez Díaz. Tengo el honor de ser presidente de la Sociedad Española de Neurología, además de haber ocupado la vicepresidencia del Consejo Español del Cerebro y de ser Fellow de la European Academy of Neurology.A lo largo de mi trayectoria, he formado parte de la junta directiva de la Sociedad Española de Neurología como vocal de comunicación, relaciones internacionales, director de cultura y vicepresidente de relaciones institucionales. También dirigí la Fundación del Cerebro.Impulsé la creación del grupo de neurooftalmología de la SEN y he formado parte de las juntas de los grupos de cefalea y neurooftalmología. Además, he sido profesor de Neurología en la Universidad Complutense de Madrid durante más de 16 años.