Las neuronas, las unidades fundamentales de nuestro sistema nervioso, se comunican constantemente a través de señales eléctricas y químicas. Esta comunicación es un delicado equilibrio entre excitación e inhibición. La inhibición es crucial para modular la actividad neuronal, dar forma a las respuestas y permitir funciones complejas como el procesamiento sensorial y la cognición. Existen diferentes mecanismos por los cuales una neurona puede ser inhibida, y dos de los más estudiados son la inhibición por hiperpolarización y la inhibición por shunting. Si bien ambos reducen la probabilidad de que una neurona dispare un potencial de acción, lo hacen de maneras fundamentalmente diferentes, lo que tiene profundas implicaciones para el procesamiento de la información neuronal.

¿Qué es la Inhibición por Shunting?
El término "shunting" (derivado del inglés, que significa desvío o derivación) se refiere a un efecto específico de la inhibición sináptica. En el contexto neuronal, la inhibición por shunting ocurre cuando un canal iónico, a menudo activado por un neurotransmisor como el GABA, se abre y aumenta la conductancia de la membrana neuronal. Si el potencial de inversión de este canal (el potencial de membrana al cual el flujo neto de iones a través del canal es cero) está cerca o ligeramente por debajo del potencial de reposo de la neurona, la apertura de estos canales no causará una hiperpolarización significativa (es decir, no hará el potencial de membrana mucho más negativo). Sin embargo, sí disminuirá drásticamente la resistencia de entrada de la neurona.
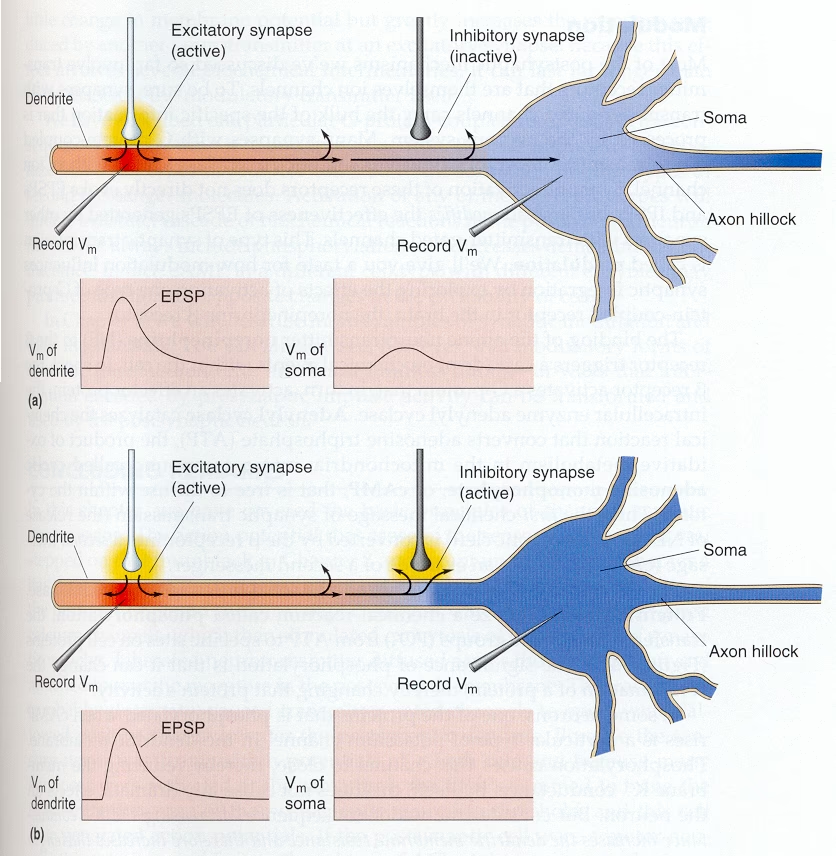
¿Cómo afecta esto a la neurona? Imagina que la neurona es un circuito eléctrico simple. La resistencia de entrada determina cuánto cambia el voltaje (potencial de membrana) en respuesta a una corriente entrante (una señal excitatoria). Al disminuir la resistencia de entrada, la inhibición por shunting reduce la magnitud del cambio de voltaje causado por una entrada excitatoria concurrente. Es como abrir una 'válvula de escape' que permite que la corriente excitatoria 'se escape' o 'se desvíe' a través de los canales abiertos, en lugar de acumularse y despolarizar la membrana hacia el umbral de disparo. Por lo tanto, aunque el potencial de membrana en reposo pueda verse poco afectado, la capacidad de la neurona para ser excitada se reduce significativamente.
Históricamente, la inhibición por shunting ha sido un candidato destacado para el mecanismo subyacente al control de ganancia en las neuronas. El control de ganancia se refiere a la capacidad de modular la sensibilidad de una neurona a sus entradas, es decir, cambiar la pendiente de su curva de entrada-salida (por ejemplo, la relación entre la fuerza de la entrada sináptica y la tasa de disparo). Se pensaba que el shunting, al reducir la efectividad de todas las entradas excitatorias de manera proporcional, actuaba como un proceso divisivo, escalando hacia abajo la respuesta neuronal.
La Inhibición por Hiperpolarización
En contraste, la inhibición por hiperpolarización ocurre cuando la apertura de canales iónicos, a menudo activados también por GABA o glicina, permite el flujo de iones (como Cl- o K+) que hacen que el potencial de membrana se vuelva más negativo que el potencial de reposo. Esto aumenta la distancia eléctrica que una entrada excitatoria debe superar para llevar el potencial de membrana hasta el umbral de disparo. La hiperpolarización tiene un efecto más directo y 'sustractivo' sobre la despolarización causada por la excitación concurrente. Esencialmente, resta un valor fijo del potencial de membrana, independientemente de la fuerza de la entrada excitatoria.
Shunting vs. Hiperpolarización: La Diferencia Clave
La distinción teórica fundamental entre la inhibición por shunting y la hiperpolarización reside en su impacto en la curva de entrada-salida de la neurona. La hiperpolarización, al hacer el potencial de membrana más negativo, típicamente desplaza la curva de entrada-salida hacia la derecha; se necesita una entrada excitatoria más fuerte para alcanzar el umbral. Este es un efecto sustractivo. Por otro lado, el shunting, al reducir la resistencia de entrada y desviar la corriente, se ha teorizado que cambia la pendiente de la curva de entrada-salida; una entrada dada produce una respuesta proporcionalmente menor. Este sería un efecto divisivo. El control de ganancia, que modula la pendiente de la curva de entrada-salida, es de vital importancia para funciones sensoriales, cognitivas y motoras normales.
Sin embargo, la realidad de cómo las neuronas integran las señales es más compleja, especialmente cuando consideramos la tasa de disparo (la frecuencia con la que una neurona genera potenciales de acción) en lugar del simple potencial de membrana. Estudios de modelado iniciales predijeron que la inhibición por shunting modularía la tasa de disparo de una manera puramente sustractiva, similar a la hiperpolarización, lo que contradecía la intuición inicial sobre el control de ganancia. Esta discrepancia generó un debate significativo en el campo.
| Característica | Inhibición por Shunting | Inhibición por Hiperpolarización |
|---|---|---|
| Efecto principal sobre Vm | Poco efecto directo en reposo (si el potencial de inversión está cerca del reposo) | Hace Vm más negativo (hiperpolarización) |
| Efecto sobre la Resistencia de Entrada | Disminuye significativamente | Generalmente aumenta (aunque depende de qué canales se abran) |
| Efecto teórico sobre Despolarización | Divisivo (reduce proporcionalmente) | Sustractivo (resta una cantidad fija) |
| Impacto en la Curva Entrada-Salida (Vm) | Cambia la pendiente | Desplaza la curva |
| Impacto en la Curva Entrada-Salida (Tasa de Disparo, condiciones simplificadas) | Sustractivo (históricamente predicho) | Sustractivo |
| Impacto en la Curva Entrada-Salida (Tasa de Disparo, condiciones realistas) | Mixto (divisivo y sustractivo), con predominancia divisiva bajo ciertas condiciones | Sustractivo |
Factores Cruciales: Ruido Sináptico y Saturación Dendrítica
La investigación más reciente ha reconciliado en parte las predicciones teóricas con las observaciones experimentales en neuronas reales. La clave para entender cómo la inhibición por shunting puede efectivamente mediar el control de ganancia de la tasa de disparo bajo condiciones fisiológicas reside en dos factores que a menudo se pasaron por alto en los modelos simplificados: el ruido sináptico y la saturación dendrítica.
Las neuronas en el cerebro intacto no reciben entradas sinápticas constantes, sino que son bombardeadas por un flujo ruidoso y fluctuante de actividad excitatoria e inhibitoria. Este ruido sináptico causa fluctuaciones irregulares en el potencial de membrana somático. Bajo estas condiciones ruidosas, el disparo neuronal se vuelve probabilístico; la neurona puede disparar incluso cuando el potencial de membrana promedio está por debajo del umbral, si una fluctuación ruidosa momentáneamente lo empuja por encima. Este 'disparo probabilístico' o 'disparo impulsado por ruido' suaviza la no linealidad del umbral de disparo. En lugar de un salto abrupto de no disparar a disparar constantemente una vez que el promedio cruza el umbral, la tasa de disparo aumenta gradualmente a medida que aumenta la entrada promedio.
El shunting, al reducir la resistencia de entrada, también reduce la amplitud de estas fluctuaciones ruidosas del potencial de membrana. Al interactuar con el umbral de disparo suavizado por el ruido, esta reducción de las fluctuaciones permite que el shunting tenga un efecto modulador sobre la tasa de disparo. Sin embargo, solo el ruido por sí solo solo permite una reducción divisiva modesta de la tasa de disparo.
Aquí es donde entra la saturación dendrítica. La mayoría de las sinapsis excitatorias se encuentran en las dendritas, lejos del soma donde se inician los potenciales de acción. A medida que la excitación dendrítica se vuelve muy fuerte, las dendritas se despolarizan significativamente. Esto reduce la fuerza impulsora para la corriente excitatoria (Vm - E_exc), un fenómeno conocido como saturación dendrítica. Además, un alto nivel de actividad sináptica cambia la estructura electrotónica de la neurona, aumentando la atenuación de las señales a medida que viajan hacia el soma. El resultado es una relación fuertemente no lineal entre la excitación dendrítica y la despolarización somática. Se necesita una excitación dendrítica muy grande para lograr incluso una despolarización somática moderada, y esta relación se vuelve menos eficiente a medida que aumenta la excitación.
La investigación ha demostrado que la combinación del suavizado del umbral por el ruido sináptico y la no linealidad causada por la saturación dendrítica es lo que permite que la inhibición por shunting ejerza un fuerte efecto predominantemente divisivo sobre la tasa de disparo en condiciones realistas. El shunting actúa sobre la relación no lineal entre la entrada excitatoria y la despolarización, y esta interacción, amplificada por el umbral suavizado, resulta en una reducción significativa de la pendiente en la curva de relación entre la entrada excitatoria y la tasa de disparo de la neurona. Ambos factores, el ruido inherente a la actividad sináptica y las propiedades no lineales de las dendritas, son cruciales para que el shunting funcione eficazmente como un mecanismo de control de ganancia en las neuronas del cerebro.
Un Concepto Diferente: El Shunt del GABA Metabólico
Es importante no confundir el efecto de inhibición neuronal por shunting, que es un fenómeno bioeléctrico relacionado con la conductancia de membrana, con el "shunt del GABA", que es una vía metabólica. Aunque el neurotransmisor GABA es a menudo el mediador de la inhibición por shunting en el cerebro, el shunt del GABA se refiere al ciclo bioquímico responsable de la síntesis, conservación y metabolismo del propio GABA.
El shunt del GABA es un proceso de circuito cerrado que comienza con el alfa-cetoglutarato, un intermediario del ciclo de Krebs (el ciclo energético celular). El alfa-cetoglutarato se transamina para formar ácido L-glutámico. Luego, una enzima clave llamada ácido glutámico descarboxilasa (GAD) cataliza la descarboxilación del glutamato para formar GABA. La presencia de GAD es un marcador distintivo de las neuronas GABAérgicas.
Para conservar el suministro de GABA, éste es metabolizado por la enzima GABA alfa-oxoglutarato transaminasa (GABA-T) de nuevo a succínico semialdehído, transfiriendo su grupo amino de vuelta al alfa-cetoglutarato para regenerar glutamato. El succínico semialdehído es luego oxidado por la succínico semialdehído deshidrogenasa (SSADH) a ácido succínico, que puede reingresar al ciclo de Krebs, completando así el bucle metabólico.
Tras ser liberado en la sinapsis, la acción del GABA finaliza principalmente por recaptación en las terminales nerviosas presinápticas y en las células gliales circundantes. Esta recaptación es mediada por transportadores de membrana específicos que requieren la presencia de iones Na+ y Cl-. El GABA recaptado puede ser reutilizado, pero el GABA que entra en las células gliales es metabolizado y no puede resintetizarse allí (ya que las células gliales carecen de GAD). Sin embargo, puede ser recuperado indirectamente a través de la glía convirtiéndolo en glutamina, que se transfiere de vuelta a la neurona, donde se convierte de nuevo en glutamato y reingresa al shunt del GABA.
Este shunt metabólico asegura que el GABA, un neurotransmisor crucial presente en altas concentraciones en muchas regiones cerebrales, sea sintetizado, utilizado y reciclado eficientemente, manteniendo así el suministro necesario para la neurotransmisión inhibitoria, incluida la que media el efecto shunting bioeléctrico.
¿Por Qué Es Importante Comprender Esto?
La comprensión detallada de cómo funcionan los diferentes tipos de inhibición, especialmente el efecto shunting bajo condiciones fisiológicas realistas, es fundamental para desentrañar los principios de la computación neuronal. El control de ganancia neuronal, mediado en gran parte por la inhibición (incluido el shunting), permite a las neuronas ajustar su sensibilidad a las entradas. Esto es vital para procesar información sensorial de manera eficiente (por ejemplo, adaptarse a diferentes niveles de contraste visual), integrar señales complejas en la toma de decisiones y mantener la estabilidad de los circuitos neuronales. Las disfunciones en estos mecanismos inhibitorios se han relacionado con diversas patologías neurológicas y psiquiátricas.
Preguntas Frecuentes
¿El efecto shunting es siempre puramente divisivo en la tasa de disparo neuronal?
No, aunque teóricamente se describió como divisivo, estudios más recientes en condiciones realistas (con ruido sináptico y saturación dendrítica) muestran que tiene componentes tanto divisivos como sustractivos en la tasa de disparo. La capacidad de actuar predominantemente de manera divisiva depende en gran medida de la presencia de ruido sináptico y la no linealidad de la saturación dendrítica.
¿Cómo se relaciona el 'shunt del GABA' con la 'inhibición por shunting'?
El 'shunt del GABA' es una vía metabólica que sintetiza y recicla el neurotransmisor GABA. La 'inhibición por shunting' es un efecto bioeléctrico en la neurona postsináptica, a menudo mediado por receptores que se activan con GABA, que reduce la resistencia de entrada y la excitabilidad de la neurona. Son conceptos diferentes, aunque el GABA producido por el shunt metabólico es el 'material' que permite la inhibición por shunting eléctrica.
¿Qué es el control de ganancia en neurociencia?
El control de ganancia es la modulación de la capacidad de respuesta de una neurona a una entrada, observado como un cambio en la pendiente de la curva de entrada-salida (por ejemplo, la relación entre la intensidad de la entrada y la frecuencia de disparo). Permite a las neuronas ajustar su sensibilidad y es crucial para el procesamiento eficiente de la información.
En resumen, el efecto shunting es un mecanismo de inhibición neuronal sutil pero poderoso que, al disminuir la resistencia de entrada, modula la forma en que una neurona integra las señales excitatorias. Contrariamente a las simplificaciones iniciales, su impacto real en la tasa de disparo, particularmente su capacidad para mediar el control de ganancia divisivo, depende críticamente de las condiciones dinámicas del entorno sináptico, como el ruido y la saturación dendrítica. Paralelamente, el shunt del GABA es una vía metabólica vital que mantiene el suministro de este importante neurotransmisor.
Si quieres conocer otros artículos parecidos a El Efecto Shunting en Neurociencia puedes visitar la categoría Neurociencia.