La memoria de trabajo es un concepto fundamental en la neurociencia cognitiva que describe un mecanismo de almacenamiento activo a corto plazo, crucial para una amplia gama de actividades cognitivas superiores como el pensamiento, el razonamiento, la planificación, la toma de decisiones y la comprensión del lenguaje. A menudo diferenciada de la memoria a corto plazo simple, la memoria de trabajo implica no solo el almacenamiento temporal de información, sino también su manipulación activa para lograr una meta actual. Es, en esencia, el espacio de trabajo mental donde procesamos la información relevante en un momento dado.

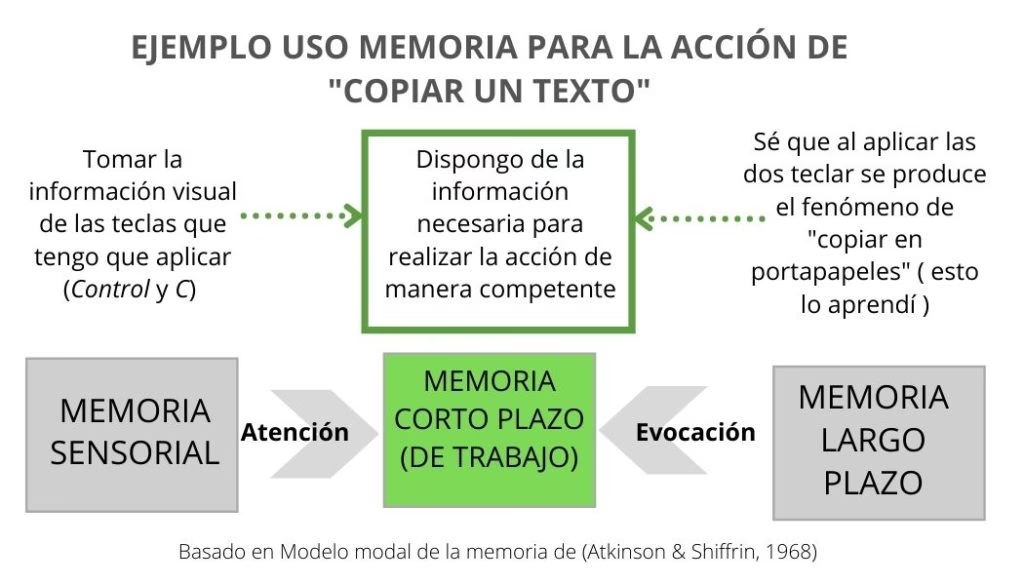
El concepto de memoria de trabajo ha evolucionado a lo largo del tiempo. Inicialmente, modelos de memoria a corto plazo, como el propuesto por Atkinson y Shiffrin en 1968, ya sugerían que esta memoria temporal actuaba como una 'memoria de trabajo' durante diversas actividades cognitivas. Sin embargo, fue Baddeley y Hitch en 1974 quienes propusieron formalmente el concepto de memoria de trabajo como un sistema cerebral dedicado al almacenamiento temporal y la manipulación de información necesaria para tareas cognitivas complejas. Más tarde, en 1999, Baddeley y Logie la describieron como el mecanismo que nos permite comprender nuestro entorno inmediato, retener experiencias recientes, adquirir nuevo conocimiento, resolver problemas y actuar sobre metas actuales.
- Modelos de Memoria de Trabajo
- La Corteza Prefrontal: Sede de la Memoria de Trabajo
- Estudio de la Memoria de Trabajo en Animales: Tareas de Retraso
- Correlatos Neuronales: La Actividad de Período de Demora
- Mecanismos Neuronales del Ejecutivo Central
- Tabla Comparativa: Memoria de Trabajo vs. Memoria de Referencia
- Preguntas Frecuentes sobre la Memoria de Trabajo
Modelos de Memoria de Trabajo
Se han propuesto varios modelos para explicar la memoria de trabajo, siendo el más influyente el de Baddeley y Hitch. El modelo original de Baddeley (1974, 1986) incluía un componente maestro, el ejecutivo central, y dos componentes 'esclavos': el bucle fonológico y el cuaderno visuoespacial. El bucle fonológico se encarga del procesamiento y almacenamiento temporal de información basada en el habla, aunque se enfatiza más el almacenamiento que el ensayo subvocal. Por otro lado, el cuaderno visuoespacial maneja la información visual y espacial que no puede ser procesada por el lenguaje, permitiendo el almacenamiento temporal de imágenes visuales y datos espaciales.
En el año 2000, Baddeley revisó su modelo para incluir un tercer componente esclavo: el buffer episódico. Este buffer de almacenamiento temporal tiene una capacidad limitada y funciona para integrar información de diversas fuentes, incluida la memoria a largo plazo. Se asume que el buffer episódico contiene episodios o fragmentos integrados y no solo actúa como un puente entre el bucle fonológico y el cuaderno visuoespacial, sino que también vincula la memoria de trabajo con la percepción y la memoria a largo plazo.
Así, los tres componentes esclavos se dedican al almacenamiento temporal de información específica: el bucle fonológico para información verbal, el cuaderno visuoespacial para información visual y espacial, y el buffer episódico para episodios integrados. El ejecutivo central, según Baddeley, es el componente maestro responsable de coordinar e integrar las operaciones de estos tres subsistemas para lograr una meta actual. Se le considera un sistema para asignar un recurso de memoria de capacidad limitada a cada componente esclavo según sus demandas. Si una tarea visuoespacial requiere más recursos que una lingüística, se asignará más capacidad a la primera. Esto permite la selección de un proceso de control o estrategia apropiada para completar una tarea. Aunque Baddeley propuso una revisión más reciente en 2012, la estructura básica del modelo se mantuvo.
La Corteza Prefrontal: Sede de la Memoria de Trabajo
La corteza prefrontal (CPF) es una región cerebral que participa en una amplia variedad de funciones cognitivas superiores, como el pensamiento, el razonamiento, la planificación y la toma de decisiones. Es considerada esencial para la inteligencia y la creatividad humanas. Varias líneas de evidencia respaldan esta idea. En humanos, la CPF constituye el 29% de la corteza cerebral, el porcentaje más alto entre los primates y mamíferos. Su maduración es más lenta que la de otras cortezas de asociación, lo que sugiere su participación en funciones complejas e integradas, más allá del procesamiento sensorial o motor básico. La CPF recibe información sensorial de cortezas de asociación, no directamente de las primarias, y envía información motora a áreas de asociación motora. El daño a la CPF en humanos se asocia con déficits en el juicio, la planificación y la toma de decisiones.
El concepto de memoria de trabajo se ha convertido en el marco más importante para interpretar y comprender las funciones de la CPF, tanto en humanos como en animales. Goldman-Rakic (1986) propuso que los déficits prefrontales observados en estudios con monos y en la clínica humana podían explicarse mediante un concepto común de memoria de trabajo (originalmente lo llamó 'memoria representacional'). Posteriormente, utilizó el término memoria de trabajo para describir la capacidad de retener información temporalmente en la mente para la comprensión, el pensamiento y la planificación. Esta idea ha sido ampliamente apoyada por numerosos estudios de neuroimagen en humanos y estudios en animales.
Estudio de la Memoria de Trabajo en Animales: Tareas de Retraso
Para comprender los mecanismos neuronales que sustentan la memoria de trabajo, son necesarios paradigmas conductuales y modelos animales adecuados. Aunque el modelo de Baddeley es útil para estudios psicológicos y neuropsicológicos humanos, adaptarlo a estudios animales es difícil. En estudios animales, la memoria de trabajo se ha definido a menudo en contraste con la 'memoria de referencia'. La memoria de referencia se refiere a la información útil a lo largo de muchos ensayos (aprendizaje de asociaciones estímulo-respuesta a largo plazo), mientras que la memoria de trabajo retiene información necesaria solo para el ensayo actual.
Las tareas de retraso (delayed tasks) han sido cruciales para examinar las funciones de la CPF en animales desde los trabajos pioneros de Jacobsen (1936), quien mostró que las lesiones bilaterales de la CPF en monos Rhesus causaban un grave y duradero deterioro en el rendimiento de la tarea de respuesta retardada (delayed-response task). Esta tarea se convirtió en un pilar experimental.
En una tarea de respuesta retardada espacial, por ejemplo, se esconde una recompensa en uno de varios lugares. Tras un período de retraso sin ver la recompensa, el sujeto debe elegir el lugar correcto basándose únicamente en la memoria de su ubicación. La posición cambia aleatoriamente en cada ensayo, exigiendo mantener activamente la información espacial específica. La tarea de alternancia retardada es similar, requiriendo recordar la elección del ensayo anterior para elegir la opuesta.
Otras tareas de memoria de trabajo no espacial incluyen la tarea de igualación a la muestra retardada (delayed matching-to-sample) y no igualación a la muestra retardada (delayed non-matching-to-sample). Aquí, se presenta un estímulo (forma, color, objeto) y, tras un retraso, se presenta junto a otro. El sujeto debe elegir el estímulo original (igualación) o el nuevo (no igualación). La información mantenida es no espacial y útil solo para el ensayo actual.
La tarea de respuesta retardada presenta varias ventajas para estudiar los mecanismos neuronales: solo se necesita memorizar una posición espacial simple, la posición del estímulo es fácil de controlar y aleatorizar, y la respuesta (manual o ocular) es fácil de cuantificar. Además, excluye el efecto de familiaridad, que puede influir en tareas de memoria de reconocimiento (donde se elige un estímulo familiar presentado previamente).
Correlatos Neuronales: La Actividad de Período de Demora
Si el deterioro de la memoria a corto plazo es una causa principal de los déficits en la tarea de respuesta retardada tras lesiones prefrontales, debe existir un mecanismo neuronal en la CPF dorsolateral responsable de este almacenamiento temporal de información espacial. Los estudios neurofisiológicos en esta área han buscado identificar estos correlatos neuronales.
Kubota y Niki (1971) y Fuster y Alexander (1971) fueron pioneros en registrar la actividad de neuronas individuales en la CPF de monos durante tareas de retraso. Fuster (1973) observó que muchas neuronas prefrontales mostraban una activación tónica sostenida durante el período de retraso, sugiriendo que esta actividad era un correlato neuronal de los procesos mnésicos a corto plazo, mientras que la actividad transitoria durante la presentación del estímulo o la respuesta se relacionaba con procesos sensoriales y motores.
Sin embargo, para que esta actividad sostenida sea un correlato de la memoria de trabajo espacial, debe mostrar diferentes patrones de activación dependiendo de la información que se mantiene (ej. la posición de la recompensa). Niki (1974) fue el primero en encontrar estos patrones diferenciales. Observó que algunas neuronas prefrontales exhibían actividad de período de demora de diferente magnitud según la posición del estímulo visual. Esta actividad, aunque en una pequeña proporción de neuronas inicialmente, fue el primer correlato neuronal identificado para el mantenimiento de información espacial.
Estudios posteriores confirmaron esta actividad de período de demora selectiva direccionalmente en varias tareas de retraso manual. Para mejorar el control experimental, se desarrolló la tarea de respuesta ocular retardada (ODR task), donde se inmoviliza la cabeza y se controla la fijación ocular durante el retraso. Esto permite presentar estímulos en múltiples posiciones del campo visual y analizar cuantitativamente los movimientos oculares.
En la tarea ODR, la actividad de período de demora en muchas neuronas de la CPF dorsolateral mostró excitación o supresión tónica sostenida, presente solo en ensayos correctos. La mayoría de esta actividad (alrededor del 80%) exhibía selectividad direccional, activándose preferentemente cuando los estímulos se presentaban en una región particular del campo visual. Esto sugirió que las neuronas prefrontales con actividad de período de demora direccional tienen 'campos de memoria' (memory fields) dentro del campo visual, análogos a los campos receptivos visuales. Estudios posteriores con múltiples posiciones de estímulo confirmaron que la mayoría de la actividad de período de demora codificaba la posición del estímulo visual (información retrospectiva), mientras que una minoría codificaba la dirección de la respuesta (información prospectiva).
La actividad de período de demora no se limita a la CPF. Se ha observado en otras áreas cerebrales como la corteza parietal, temporal, somatosensorial, premotora, visual primaria, colículo superior, ganglios basales, hipocampo, tálamo e incluso la médula espinal. Sin embargo, la información codificada por esta actividad varía entre áreas. Mientras que en la CPF predomina la codificación de la ubicación del estímulo visual, en la corteza parietal y el núcleo mediodorsal del tálamo tiende a codificar la dirección de la respuesta sacádica.
Además, la información codificada por la actividad de período de demora en la CPF también depende de los requisitos de la tarea. En la tarea de respuesta retardada codifica principalmente la posición espacial; en la tarea de igualación a la muestra retardada, puede codificar características físicas no espaciales (color, forma). También se ha reportado actividad de período de demora en la CPF que representa información táctil, auditiva, reglas de tarea, diferencias entre tareas, recompensa esperada, cantidad numérica, distancia relativa entre estímulos, sincronización temporal y orden temporal de los estímulos.
Mecanismos Neuronales del Ejecutivo Central
Aunque el modelo de Baddeley es abstracto, el ejecutivo central se considera el componente que coordina y controla las operaciones de múltiples sistemas cognitivos, integrando señales de 'arriba hacia abajo' (top-down) y de 'abajo hacia arriba' (bottom-up) para lograr una meta. Estas funciones se alinean con las atribuidas a la CPF, que recibe información diversa y controla otras áreas corticales y subcorticales. Por ello, se cree que el ejecutivo central refleja las funciones de la CPF, y el término 'función ejecutiva' se usa a menudo para describir las funciones prefrontales.
Una forma de examinar las funciones del ejecutivo central es mediante paradigmas de doble tarea (dual-task paradigm), donde se requiere realizar dos tareas simultáneamente. Bajo esta condición, el rendimiento suele empeorar en una o ambas tareas ('interferencia de doble tarea'). Esto se atribuye a una capacidad limitada de recurso cognitivo, una capacidad general de procesamiento de información compartida por tareas concurrentes. El ejecutivo central, en el modelo de Baddeley, sería el sistema que asigna este recurso limitado a cada tarea según su demanda.
Watanabe y Funahashi (2014, 2015) investigaron esto neurofisiológicamente en monos usando una tarea de memoria de trabajo espacial y una tarea de atención espacial bajo condiciones de tarea única y doble tarea. Ambas tareas reclutaron poblaciones neuronales superpuestas en la CPF dorsolateral. Observaron que la interferencia conductual aumentaba con la dificultad de la tarea concurrente, y este efecto se correlacionó con el grado de solapamiento de las poblaciones neuronales reclutadas. Crucialmente, la magnitud de la actividad de período de demora que representaba la información espacial de la tarea de memoria se atenuó significativamente en la condición de doble tarea en comparación con la tarea única, y esta atenuación fue mayor al aumentar la dificultad de la tarea de atención.
Estos hallazgos sugieren que la capacidad de una población neuronal prefrontal para representar información relevante para la tarea disminuye en proporción al aumento de la demanda de la tarea concurrente. La interferencia de doble tarea parece originarse en el reclutamiento competitivo y sobrecargado de poblaciones neuronales comunes en la CPF por las dos tareas. La capacidad de procesamiento de información de una sola neurona parece ser limitada, pero puede asignarse o reasignarse flexiblemente entre tareas concurrentes según sus necesidades, mejorando el rendimiento conductual cuando se aumenta su asignación a una tarea.
La atenuación y posterior 'reactivación' (reawakening) de la actividad neuronal observada en la condición de doble tarea sugiere que el recurso cognitivo en la CPF se divide y asigna a las tareas según sus demandas. Esto apoya la idea de que la capacidad limitada de los recursos cognitivos es resultado de la capacidad computacional limitada de las neuronas individuales. Aunque los mecanismos exactos de esta división y asignación de recursos en la CPF aún no están claros, su identificación es fundamental para comprender el control ejecutivo de la CPF y el sistema neural responsable del ejecutivo central en el modelo de memoria de trabajo.
Tabla Comparativa: Memoria de Trabajo vs. Memoria de Referencia
| Característica | Memoria de Trabajo | Memoria de Referencia |
|---|---|---|
| Utilidad de la Información | Necesaria solo para el ensayo/situación actual. | Útil a lo largo de muchos ensayos o experimentos. |
| Naturaleza de la Información | Varía de ensayo a ensayo (ej. posición, objeto, regla). | Asociación fija entre un estímulo y una respuesta. |
| Mecanismo Clave | Mantenimiento activo y manipulación temporal de información. | Formación y mantenimiento a largo plazo de asociaciones. |
| Tareas Típicas | Tareas de respuesta retardada, alternancia retardada, igualación/no igualación a la muestra retardada. | Tareas de discriminación visual. |
| Región Cerebral Clave (Animales) | Corteza Prefrontal Dorsolateral. | Hipocampo, Lóbulo Temporal Medial. |
Preguntas Frecuentes sobre la Memoria de Trabajo
¿Qué diferencia hay entre memoria de trabajo y memoria a corto plazo?
Aunque a veces se usan indistintamente, la memoria a corto plazo se refiere más a la capacidad simple de almacenamiento temporal pasivo. La memoria de trabajo es un concepto más amplio que incluye no solo el almacenamiento temporal sino también la manipulación activa y el procesamiento de esa información para realizar tareas cognitivas.
¿Dónde se localiza la memoria de trabajo en el cerebro?
La corteza prefrontal, especialmente la corteza prefrontal dorsolateral, es considerada la región clave para la memoria de trabajo, particularmente para las funciones del ejecutivo central y el mantenimiento activo de información. Sin embargo, la memoria de trabajo involucra una red de áreas cerebrales, incluyendo cortezas parietales y temporales, que colaboran en el procesamiento y almacenamiento de diferentes tipos de información.
¿Cómo se estudia la memoria de trabajo en neurociencia?
Se estudia utilizando tareas conductuales que requieren el mantenimiento y/o manipulación de información durante un breve período, como las tareas de respuesta retardada o de doble tarea. En animales, se combinan con técnicas neurofisiológicas (registro de actividad neuronal) o estudios de lesión. En humanos, se usan tareas conductuales, neuroimagen (fMRI, EEG) y estudios de pacientes con daño cerebral.
¿Qué es la actividad de período de demora?
Es la actividad neuronal sostenida que se observa en ciertas neuronas de áreas como la corteza prefrontal durante el intervalo de tiempo entre la presentación de un estímulo relevante y la respuesta requerida. Se considera un correlato neural clave del mecanismo por el cual se mantiene activa la información en la memoria de trabajo.
¿Qué ocurre si la memoria de trabajo está afectada?
El daño a la corteza prefrontal u otras áreas de la red de memoria de trabajo puede causar déficits significativos en las funciones ejecutivas, incluyendo problemas para mantener la concentración, planificar, resolver problemas, tomar decisiones, seguir instrucciones complejas y resistir distracciones (lo que se relaciona con la interferencia de doble tarea).
La memoria de trabajo es, por lo tanto, un sistema cognitivo dinámico y fundamental, cuya comprensión a nivel neuronal, especialmente en la corteza prefrontal, sigue siendo un área activa y fascinante de investigación en neurociencia. Los estudios continúan desvelando cómo el cerebro mantiene, manipula y controla la información que necesitamos 'tener en mente' para navegar por nuestro complejo mundo.
Si quieres conocer otros artículos parecidos a Memoria de Trabajo: El Director Cognitivo puedes visitar la categoría Neurociencia.