El funcionamiento de nuestro sistema nervioso, la capacidad de nuestros músculos para contraerse y, en esencia, la vida misma, dependen de una propiedad fundamental de las células: el potencial de membrana. Esta diferencia de voltaje a través de la delgada barrera que separa el interior del exterior celular es el motor de la excitabilidad. Durante mucho tiempo, la naturaleza exacta y la magnitud de este potencial, especialmente el potencial de membrana en reposo, fueron un misterio. Se sabía que las células excitables estaban polarizadas, siendo el interior negativo con respecto al exterior, y se observaba una distribución iónica desigual: altas concentraciones de potasio (K+) dentro de la célula y altas concentraciones de sodio (Na+) y cloruro (Cl-) fuera.

La primera hipótesis cuantitativa significativa provino de Julius Bernstein en 1902. Basándose en el conocimiento de que el K+ era el principal catión intracelular, Bernstein propuso que la membrana celular en reposo era permeable principalmente al K+ e impermeable a otros iones. Si esto fuera cierto, la diferencia de potencial a través de la membrana podría explicarse por la tendencia del K+ a difundir a favor de su gradiente de concentración, generando una carga eléctrica. La herramienta matemática para describir esta situación era la Ecuación de Nernst.

- La Hipótesis de Bernstein y la Ecuación de Nernst
- La Necesidad de un Modelo Multiónico
- El Avance de Goldman: El Campo Constante
- La Ecuación de Goldman-Hodgkin-Katz (GHK) para el Potencial de Membrana
- Permeabilidad Iónica y el Potencial de Reposo
- Diferencias Clave: Nernst vs GHK
- La Relevancia y Aplicación de la Ecuación GHK
- Preguntas Frecuentes
La Hipótesis de Bernstein y la Ecuación de Nernst
La Ecuación de Nernst describe el potencial de equilibrio para un ion específico a través de una membrana permeable solo a ese ion. Este potencial de equilibrio es el voltaje transmembrana en el cual el flujo neto de un ion a través de la membrana es cero, ya que la fuerza eléctrica que mueve el ion en una dirección se equilibra exactamente con la fuerza química (dada por el gradiente de concentración) que lo mueve en la dirección opuesta.
Matemáticamente, la Ecuación de Nernst para un ion X se expresa generalmente como:
EX = (RT/zF) * ln([X]o / [X]i)
Donde EX es el potencial de equilibrio para el ion X, R es la constante de los gases, T es la temperatura absoluta, z es la valencia del ion, F es la constante de Faraday, y [X]o y [X]i son las concentraciones (o más precisamente, actividades) del ion fuera y dentro de la célula, respectivamente. A temperatura ambiente y usando logaritmo base 10, esto se simplifica a aproximadamente (58/z) * log([X]o / [X]i) milivoltios.
Bernstein probó su hipótesis observando el "potencial de lesión" en músculos de rana, que era negativo en el interior, como predecía la permeabilidad al K+. También vio que aumentar el K+ externo despolarizaba la membrana, en la dirección correcta. Sin embargo, los resultados no coincidían exactamente con las predicciones de Nernst en todos los rangos de concentración de K+, y lo que es más importante, se descubrió que la membrana celular en reposo no era impermeable a otros iones.
La Necesidad de un Modelo Multiónico
Experimentos posteriores, como los de Boyle y Conway (1941) con cloruro y Levi y Ussing (1948) con sodio usando isótopos radiactivos, demostraron claramente que la membrana muscular era permeable al Cl- y al Na+, además de al K+. Cuando Hodgkin y Huxley (1939) y Curtis y Cole (1942) lograron medir directamente el potencial transmembrana en el axón gigante del calamar, y Graham y Gerard (1946) y Ling y Gerard (1949) lo hicieron en músculo de rana, confirmaron que el potencial de membrana en reposo (Er) estaba cerca del potencial de equilibrio del potasio (EK) predicho por Nernst, pero no era idéntico. Además, las variaciones en el K+ externo no siempre seguían la relación Nernstiana perfecta, especialmente a bajas concentraciones de K+. Era evidente que un modelo que considerara la permeabilidad a múltiples iones era necesario para explicar el potencial de membrana observado.
El Avance de Goldman: El Campo Constante
En 1943, David Goldman dio un paso crucial. Propuso un tratamiento matemático para la relación entre el potencial transmembrana, las concentraciones de varios iones a ambos lados de la membrana, y las movilidades iónicas. Para simplificar las complejas ecuaciones de electrodifusión (como la de Nernst-Planck) y hacerlas manejables, Goldman introdujo una suposición clave: que el gradiente de potencial eléctrico a través de la membrana era constante. Esta se conoció como la hipótesis del campo constante.

Bajo esta suposición, Goldman pudo derivar expresiones para el flujo (o densidad de corriente) de cada ion individual a través de la membrana en función del potencial transmembrana y las concentraciones iónicas a ambos lados. Estas ecuaciones describen cómo la fuerza eléctrica y la fuerza química impulsan el movimiento de cada tipo de ion permeable.
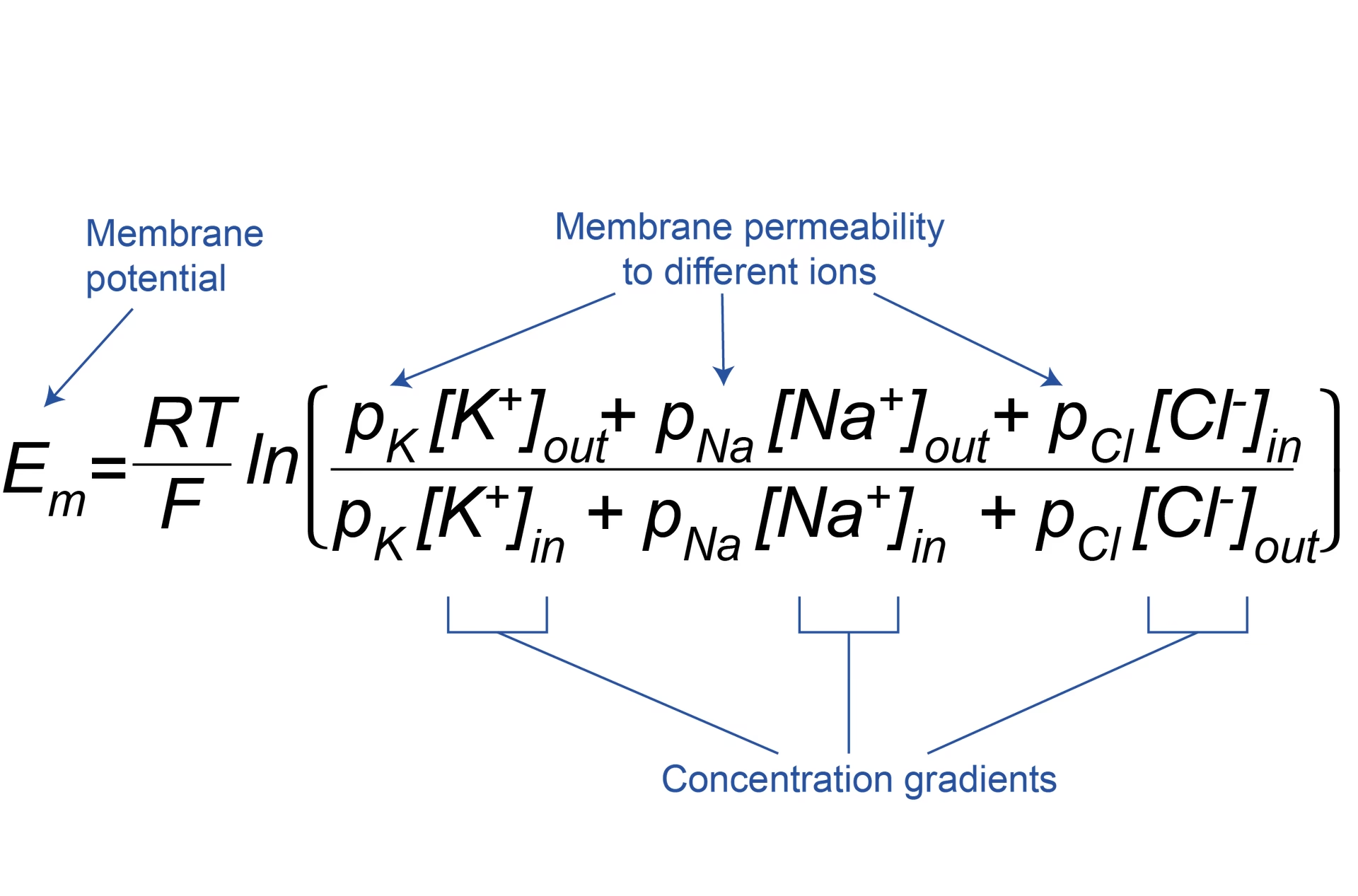
La Ecuación de Goldman-Hodgkin-Katz (GHK) para el Potencial de Membrana
El trabajo de Goldman fue fundamental, pero fue Hodgkin y Katz en 1949 quienes aplicaron y simplificaron la ecuación de Goldman para analizar datos del axón gigante del calamar. La ecuación resultante, que se conoció como la Ecuación de Goldman-Hodgkin-Katz (GHK) para el potencial de voltaje, se ha convertido en una herramienta estándar en neurofisiología para entender los potenciales de membrana en reposo y también los potenciales de acción.
La Ecuación GHK para el potencial de membrana (ψ) en un estado estacionario (donde la corriente neta total a través de la membrana es cero) que considera la permeabilidad a K+, Na+ y Cl- es:
ψ = - (RT/F) * ln ( [PK(K+)i + PNa(Na+)i + PCl(Cl-)o] / [PK(K+)o + PNa(Na+)o + PCl(Cl-)i] )
Aquí, PK, PNa y PCl son los coeficientes de permeabilidad de la membrana para el potasio, sodio y cloruro, respectivamente. Estos coeficientes reflejan la facilidad con la que cada ion puede atravesar la membrana y son cruciales porque no solo dependen de las concentraciones, sino también de las propiedades de la membrana y de los canales iónicos presentes. La ecuación muestra que el potencial de membrana no es simplemente el potencial de equilibrio de un ion, sino un promedio ponderado de los potenciales de equilibrio de los iones permeables, donde los pesos son las permeabilidades relativas de la membrana a esos iones.
La justificación para establecer la corriente neta total a cero (J=0) para derivar la ecuación GHK de voltaje es que, en el estado estacionario del potencial de reposo, la suma de las corrientes entrantes de cationes y salientes de aniones debe ser igual a la suma de las corrientes salientes de cationes y entrantes de aniones. Es decir, no hay acumulación neta de carga a través de la membrana con el tiempo.
Permeabilidad Iónica y el Potencial de Reposo
La Ecuación GHK es poderosa porque incorpora la permeabilidad de la membrana a cada ion. En la membrana neuronal o muscular en reposo, la permeabilidad al K+ (PK) es significativamente mayor que la permeabilidad al Na+ (PNa). Esto se debe a que en reposo hay muchos canales de fuga de K+ abiertos, mientras que los canales de Na+ (responsables del potencial de acción) están mayormente cerrados. La permeabilidad al Cl- (PCl) también puede ser considerable, a menudo mayor que la del K+.

En el músculo de rana, por ejemplo, se ha sugerido que el cloruro puede estar cerca de su propio equilibrio electroquímico en reposo (JCl ≈ 0), lo que significaría que el potencial de membrana en reposo es aproximadamente igual al potencial de equilibrio del cloruro (ECl). Si JCl = 0, la Ecuación GHK se simplifica y solo quedan los términos de K+ y Na+.
A pesar de su potencia, medir con precisión los coeficientes de permeabilidad puede ser difícil, ya que pueden variar con el potencial de membrana o con las concentraciones iónicas, especialmente durante la actividad eléctrica de la célula (como un potencial de acción). Sin embargo, incluso asumiendo que los valores de P son constantes en el rango del potencial de reposo, la Ecuación GHK ha permitido estimar las permeabilidades relativas. Para el músculo de rana en reposo, las estimaciones sugieren que PK es entre 20 y 100 veces mayor que PNa, y PCl podría ser mayor que PK.
La Ecuación GHK solo considera iones monovalentes como K+, Na+ y Cl- porque son los que más contribuyen al potencial de membrana en la mayoría de los casos. Para que un ion contribuya significativamente, debe cumplir tres requisitos: tener un gradiente de concentración, tener una concentración suficiente en al menos un lado de la membrana, y tener un coeficiente de permeabilidad lo suficientemente grande. Iones como el H+ o el Ca2+ pueden no cumplir estos requisitos en reposo (H+ por baja concentración, Ca2+ por baja permeabilidad en la membrana de reposo), aunque el Ca2+ juega un papel crucial en la actividad sináptica y otros procesos.
Diferencias Clave: Nernst vs GHK
Es fundamental comprender la distinción entre la Ecuación de Nernst y la Ecuación GHK:
| Característica | Ecuación de Nernst | Ecuación de Goldman-Hodgkin-Katz (GHK) |
|---|---|---|
| Ion(es) considerados | Un solo ion específico | Múltiples iones permeables (típicamente K+, Na+, Cl-) |
| Potencial calculado | Potencial de equilibrio para ese ion (EX) | Potencial de membrana en estado estacionario (ψ), a menudo el potencial de reposo |
| Suposición clave sobre el flujo | Flujo neto del ion = 0 (equilibrio electroquímico) | Corriente neta total = 0 (estado estacionario) |
| Considera permeabilidad | Implícita (membrana permeable solo a ese ion) | Explícitamente incluye coeficientes de permeabilidad (PX) para cada ion |
| Aplicabilidad | Predice el potencial si solo un ion fuera permeable. Utilizado como referencia para potenciales de equilibrio individuales. | Describe el potencial en membranas permeables a múltiples iones. Más precisa para el potencial de reposo real de las células. |
En esencia, la Ecuación de Nernst describe una situación idealizada para un solo ion en equilibrio, mientras que la Ecuación GHK describe una situación más realista para el potencial de membrana en reposo, donde varios iones se mueven a través de la membrana, pero la corriente neta total es cero debido a las permeabilidades y gradientes.
La Relevancia y Aplicación de la Ecuación GHK
La Ecuación de Goldman-Hodgkin-Katz ha demostrado ser increíblemente productiva en neurofisiología y campos relacionados. Ha permitido a los investigadores no solo comprender mejor el potencial de membrana en reposo, sino también analizar y modelar fenómenos dinámicos como los potenciales de acción, donde las permeabilidades a Na+ y K+ cambian drásticamente con el voltaje. Aunque la suposición del campo constante es una simplificación, la ecuación GHK ha sido validada por una gran cantidad de datos experimentales y sigue siendo una herramienta esencial para relacionar las concentraciones iónicas, las permeabilidades de la membrana y el potencial eléctrico transmembrana.
Preguntas Frecuentes
¿Por qué la Ecuación de Nernst no es suficiente para describir el potencial de reposo?
La Ecuación de Nernst es ideal para un solo ion. La membrana celular real en reposo es permeable a varios iones (K+, Na+, Cl-), no solo a uno. El potencial de reposo es el resultado de las corrientes de todos estos iones, no solo del punto de equilibrio de uno solo.
¿Qué significa la permeabilidad en la Ecuación GHK?
La permeabilidad (P) es un coeficiente que representa la facilidad con la que un ion específico puede cruzar la membrana. Depende de factores como el número de canales iónicos abiertos para ese ion y sus propiedades, y la solubilidad del ion en la membrana. Una mayor permeabilidad significa que un ion tiene una mayor influencia en el potencial de membrana, dado su gradiente de concentración.
¿La Ecuación GHK predice el potencial de acción?
La Ecuación GHK describe el potencial de membrana en un estado estacionario (corriente neta cero), lo cual es característico del potencial de reposo. Sin embargo, los principios subyacentes (relación entre potencial, concentraciones y permeabilidades) son aplicables. Durante un potencial de acción, las permeabilidades a Na+ y K+ cambian drásticamente y rápidamente con el voltaje. Aunque la ecuación GHK de voltaje en su forma simple no describe la dinámica del potencial de acción, las ecuaciones de flujo GHK son componentes de modelos más complejos (como el modelo de Hodgkin-Huxley) que sí describen estas dinámicas al considerar cómo las permeabilidades cambian con el tiempo y el voltaje.
¿Qué es la suposición del campo constante?
Es la suposición simplificadora de que el gradiente de potencial eléctrico (el cambio de voltaje por unidad de distancia) a través de la membrana es uniforme. Esto facilitó la integración de las ecuaciones de electrodifusión para obtener la forma manejable de la Ecuación GHK de voltaje. Aunque es una idealización, los resultados obtenidos con esta suposición se ajustan bien a muchos datos experimentales.
En resumen, la Ecuación de Goldman-Hodgkin-Katz representa un hito fundamental en nuestra comprensión de cómo los gradientes iónicos y la permeabilidad de la membrana se combinan para establecer el potencial eléctrico a través de la membrana celular, un fenómeno esencial para la función de las células excitables.
Si quieres conocer otros artículos parecidos a La Ecuación de Goldman: Clave del Potencial Neuronal puedes visitar la categoría Neurociencia.