Los canales iónicos son proteínas esenciales incrustadas en las membranas celulares, actuando como compuertas que controlan el paso de iones específicos como sodio (Na+), potasio (K+), calcio (Ca2+) y cloruro (Cl-). Este flujo de iones es fundamental para una vasta gama de procesos biológicos, desde la transmisión de señales nerviosas en el cerebro hasta la contracción muscular y la secreción hormonal. Si bien la apertura (activación) y el cierre (desactivación) de estos canales en respuesta a cambios en el voltaje de la membrana o la unión de ligandos son bien conocidos, existe otro estado crucial que regula su función: la inactivación.
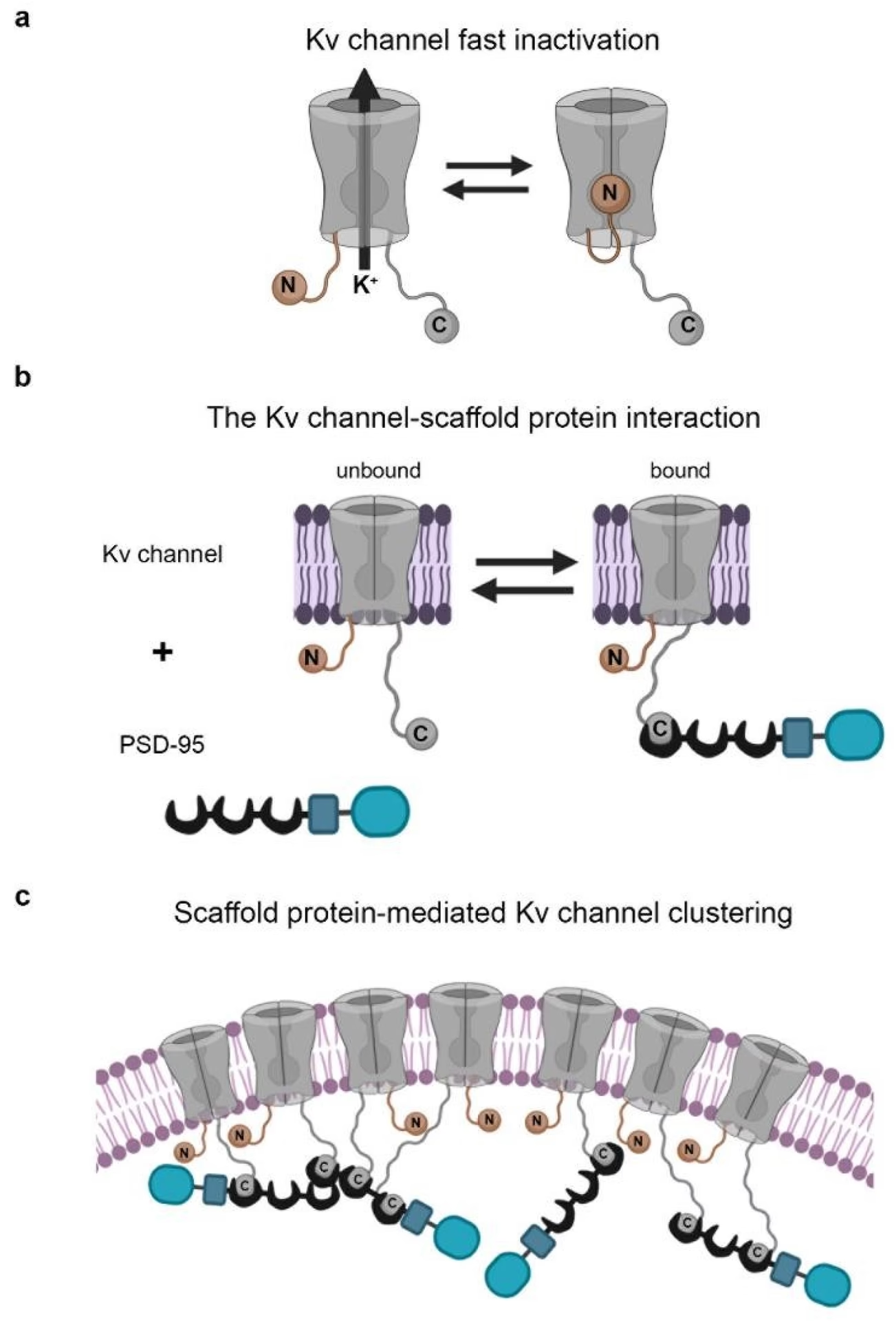
La inactivación es un proceso mediante el cual un canal iónico que se ha abierto deja de conducir iones, incluso si el estímulo que causó su apertura (como un cambio de voltaje) sigue presente. Es un mecanismo intrínseco del canal que lo pone temporalmente fuera de servicio. Esta capacidad de inactivarse es vital para controlar la duración y la frecuencia de los potenciales de acción, permitiendo que las neuronas se recuperen y puedan disparar repetidamente, lo que es fundamental para el procesamiento de información en el sistema nervioso.
¿Qué es la Inactivación Tipo N?
La inactivación tipo N es uno de los principales mecanismos de inactivación de los canales iónicos, especialmente prominente en ciertos tipos de canales de potasio dependientes de voltaje (Kv). Se caracteriza por ser un proceso rápido, que se activa poco después de que el canal se abre. Su nombre, 'tipo N', se refiere a que a menudo implica la participación del dominio N-terminal (el extremo amino) de la proteína del canal o de una subunidad asociada.
El principio fundamental detrás de la inactivación tipo N es que una parte de la propia proteína del canal actúa como un 'tapón' o 'bola' que ocluye físicamente la boca intracelular del poro del canal una vez que este se ha abierto. Esta oclusión bloquea el paso de iones, deteniendo la conducción.
El Modelo Clásico de la "Bola y Cadena"
El modelo de la bola y cadena es la descripción más estudiada y emblemática de la inactivación tipo N. Fue propuesto inicialmente para explicar la inactivación de los canales de sodio, aunque estudios posteriores mostraron que estos canales utilizan un mecanismo ligeramente diferente (una 'tapa con bisagra'). Sin embargo, el modelo de bola y cadena se demostró claramente aplicable a la inactivación rápida del canal de potasio Shaker de la mosca de la fruta, un canal modelo muy utilizado en investigación.
En el canal Shaker, la inactivación tipo N se logra mediante una "bola" peptídica compuesta por aproximadamente 19 residuos de aminoácidos. Esta bola está unida a la membrana celular por una "cadena", que es una secuencia flexible de unos 60 residuos de aminoácidos. Cuando el canal Shaker se abre en respuesta a la despolarización de la membrana, esta cadena permite que la bola peptídica se mueva libremente en el espacio intracelular cerca de la boca del poro.
De manera aleatoria, la bola puede entrar y encajar en la abertura intracelular del poro. Cuando esto sucede, la bola bloquea físicamente el paso de los iones potasio a través del canal. El canal sigue estando en un estado 'abierto' desde el punto de vista de su activación por voltaje, pero está inactivado funcionalmente porque no puede conducir iones. La conducción se detiene hasta que la bola se desprende del poro, lo que permite que el canal se recupere de la inactivación y vuelva a estar disponible para conducir iones si aún está en estado abierto o si se reabre.
Este mecanismo es análogo a poner un tapón en un desagüe: el desagüe sigue siendo un conducto, pero el tapón impide que el agua pase. La velocidad con la que ocurre esta inactivación depende en gran medida de la flexibilidad y longitud de la cadena, así como de la afinidad de la bola por su sitio de unión en el poro.
Inactivación Tipo N en Otros Canales
Si bien el modelo de bola y cadena describe el mecanismo en canales como Shaker (donde la bola puede ser el N-terminal de la subunidad principal o de una subunidad accesoria β), el concepto de inactivación tipo N se extiende a otros canales Kv. Por ejemplo, en el canal Kv1.4, el propio dominio N-terminal de la subunidad principal actúa como la 'bola' que ocluye el poro una vez que el canal se abre. Al igual que en el modelo clásico, esto requiere que el canal esté en un estado abierto para que el N-terminal acceda a su sitio de bloqueo en la boca intracelular del poro.
Es importante destacar que la inactivación tipo N en Kv1.4 parece requerir que el canal esté completamente abierto para que el dominio N-terminal se una eficazmente y bloquee el poro. Esto contrasta con otros tipos de inactivación o con la inactivación en algunos otros canales (como algunas formas de inactivación tipo N en Shaker o la inactivación tipo C en otros canales) que pueden ocurrir desde estados cerrados o parcialmente activados.
Contraste con la Inactivación Tipo C
La inactivación tipo N a menudo coexiste con otro mecanismo de inactivación, conocido como inactivación tipo C. Mientras que la inactivación tipo N es rápida y mediada por un 'tapón' intracelular, la inactivación tipo C es generalmente más lenta y parece implicar cambios conformacionales más globales en la estructura del canal, afectando tanto la cara intracelular como la extracelular.
Las diferencias clave entre estos dos mecanismos incluyen:
- Velocidad: La inactivación tipo N es rápida (milisegundos), mientras que la tipo C es lenta (decenas o cientos de milisegundos).
- Mecanismo Molecular: La tipo N implica la oclusión física del poro por un dominio N-terminal o una subunidad asociada. La tipo C parece involucrar cambios más sutiles en la conformación del poro, posiblemente relacionados con la constricción de la boca del poro o cambios en el filtro de selectividad.
- Dependencia del Estado Abierto: La inactivación tipo N (al menos en canales como Kv1.4) requiere que el canal esté en un estado abierto para que el 'tapón' acceda al poro. La inactivación tipo C puede ocurrir desde estados cerrados, parcialmente abiertos o completamente abiertos.
- Sensibilidad: La inactivación tipo N es sensible a modificaciones en el dominio N-terminal o la cadena. La inactivación tipo C es sensible a una variedad de factores, incluyendo iones extracelulares, toxinas, osmolaridad intracelular y mutaciones en diferentes partes del canal (intracelulares y extracelulares).
En canales que poseen ambos mecanismos (como Kv1.4), la inactivación tipo N domina la fase inicial de inactivación debido a su velocidad. La inactivación tipo C, al ser más lenta, a menudo determina la tasa de recuperación de la inactivación y juega un papel crucial en la disponibilidad del canal para futuras estimulaciones, afectando así las propiedades de disparo a largo plazo de la neurona.
Importancia Funcional
La presencia y las propiedades específicas de los mecanismos de inactivación tipo N y tipo C son cruciales para determinar cómo se comporta una neurona en respuesta a diferentes patrones de estimulación. La inactivación rápida tipo N limita la duración del potencial de acción y contribuye al período refractario absoluto o relativo, asegurando que la neurona no dispare demasiado rápido.
Por otro lado, la inactivación lenta tipo C influye en la adaptación de la frecuencia de disparo, permitiendo que la neurona ajuste su excitabilidad durante una estimulación prolongada. La interacción y el acoplamiento entre la activación del canal y sus estados de inactivación (tipo N y tipo C) son complejos y varían entre los diferentes tipos de canales iónicos, pero son fundamentales para la plasticidad y la función normal del sistema nervioso.
Tabla Comparativa: Inactivación Tipo N vs. Tipo C
| Característica | Inactivación Tipo N | Inactivación Tipo C |
|---|---|---|
| Velocidad | Rápida (milisegundos) | Lenta (decenas a cientos de milisegundos) |
| Mecanismo | Oclusión física del poro (dominio N-terminal o 'bola') | Cambios conformacionales globales (intracelulares y extracelulares) |
| Modelo Clásico | Bola y Cadena (canal Shaker) | Menos definido molecularmente |
| Dependencia del Estado Abierto (Ej: Kv1.4) | Requiere estado abierto (o completamente abierto) | Puede ocurrir desde estados cerrados, parcial o completamente abiertos |
| Región Clave del Canal | Dominio N-terminal / Boca intracelular del poro | Estructura del poro, filtro de selectividad, regiones intra y extracelulares |
| Sensibilidad | Mutaciones/alteraciones en el N-terminal o 'cadena' | Iones extracelulares, TEA, cambios osmóticos, mutaciones en varias regiones |
| Impacto Principal | Duración inicial del potencial de acción, periodo refractario | Recuperación de la inactivación, adaptación de la frecuencia de disparo |
Preguntas Frecuentes (FAQ)
Q: ¿Todos los canales iónicos tienen inactivación tipo N o tipo C?
A: No, la presencia y el tipo de mecanismos de inactivación varían mucho entre los diferentes tipos de canales iónicos. Algunos canales pueden tener solo un tipo de inactivación, ambos, o ninguno.
Q: ¿El mecanismo de la 'bola y cadena' es exactamente lo mismo que la inactivación tipo N?
A: El mecanismo de la 'bola y cadena' es un modelo específico y bien estudiado de inactivación tipo N, particularmente aplicable al canal Shaker. La inactivación tipo N es una categoría más amplia que se refiere a la inactivación rápida mediada por un dominio N-terminal que ocluye el poro, incluso si el mecanismo molecular exacto difiere ligeramente (como en Kv1.4 donde el propio N-terminal actúa como la 'bola').
Q: ¿Por qué es importante la inactivación para la función neuronal?
A: La inactivación es crucial para limitar la duración de los potenciales de acción, establecer periodos refractarios (momentos en que la neurona es menos propensa a disparar) y permitir que las neuronas disparen repetidamente a frecuencias controladas. Sin inactivación, las señales nerviosas serían muy diferentes y la comunicación en el cerebro se vería gravemente afectada.
Q: ¿La inactivación tipo C es solo una versión más lenta de la tipo N?
A: No, aunque ambas son formas de inactivación, sus mecanismos moleculares son distintos. La tipo N es un bloqueo físico del poro, mientras que la tipo C implica cambios conformacionales más amplios en la estructura del canal. Sus sensibilidades y dependencias del estado del canal también difieren.
Conclusión
La inactivación de los canales iónicos es un proceso dinámico y finamente regulado que añade una capa esencial de complejidad a la función de estas proteínas. Mecanismos como la inactivación tipo N, ejemplificada por el elegante modelo de la bola y cadena, y la inactivación tipo C, con sus propiedades más lentas y mecanismos conformacionales, no son meras curiosidades biológicas. Son procesos fundamentales que controlan la excitabilidad de las células, dan forma a las señales eléctricas que viajan por nuestro sistema nervioso y, en última instancia, sustentan todas las funciones cerebrales, desde el pensamiento hasta el movimiento.
Si quieres conocer otros artículos parecidos a Inactivación: Bola y Cadena y Tipo N puedes visitar la categoría Neurociencia.