Nuestro mundo está lleno de sonidos, desde el murmullo de una conversación hasta la estruendosa música de un concierto o el sutil crujido de las hojas bajo nuestros pies. Pero, ¿cómo percibe nuestro cerebro toda esta información sonora? La respuesta reside, en gran parte, en una región fascinante: el córtex auditivo. Este centro cerebral es el destino final de las señales que viajan desde nuestros oídos, transformando vibraciones mecánicas en las complejas percepciones auditivas que experimentamos.
- ¿Qué es el Córtex Auditivo y Dónde se Procesa el Sonido?
- Organización Compleja: Áreas Nucleares y de Cinturón
- Arquitectura Celular en Capas: Una Estructura Laminar Sofisticada
- La Intrincada Red de Conexiones del Córtex Auditivo
- Especialización Funcional y el Procesamiento del Sonido Complejo
- Comparativa: Áreas Nucleares (Core) vs. Áreas de Cinturón (Belt)
- Preguntas Frecuentes sobre el Córtex Auditivo
¿Qué es el Córtex Auditivo y Dónde se Procesa el Sonido?
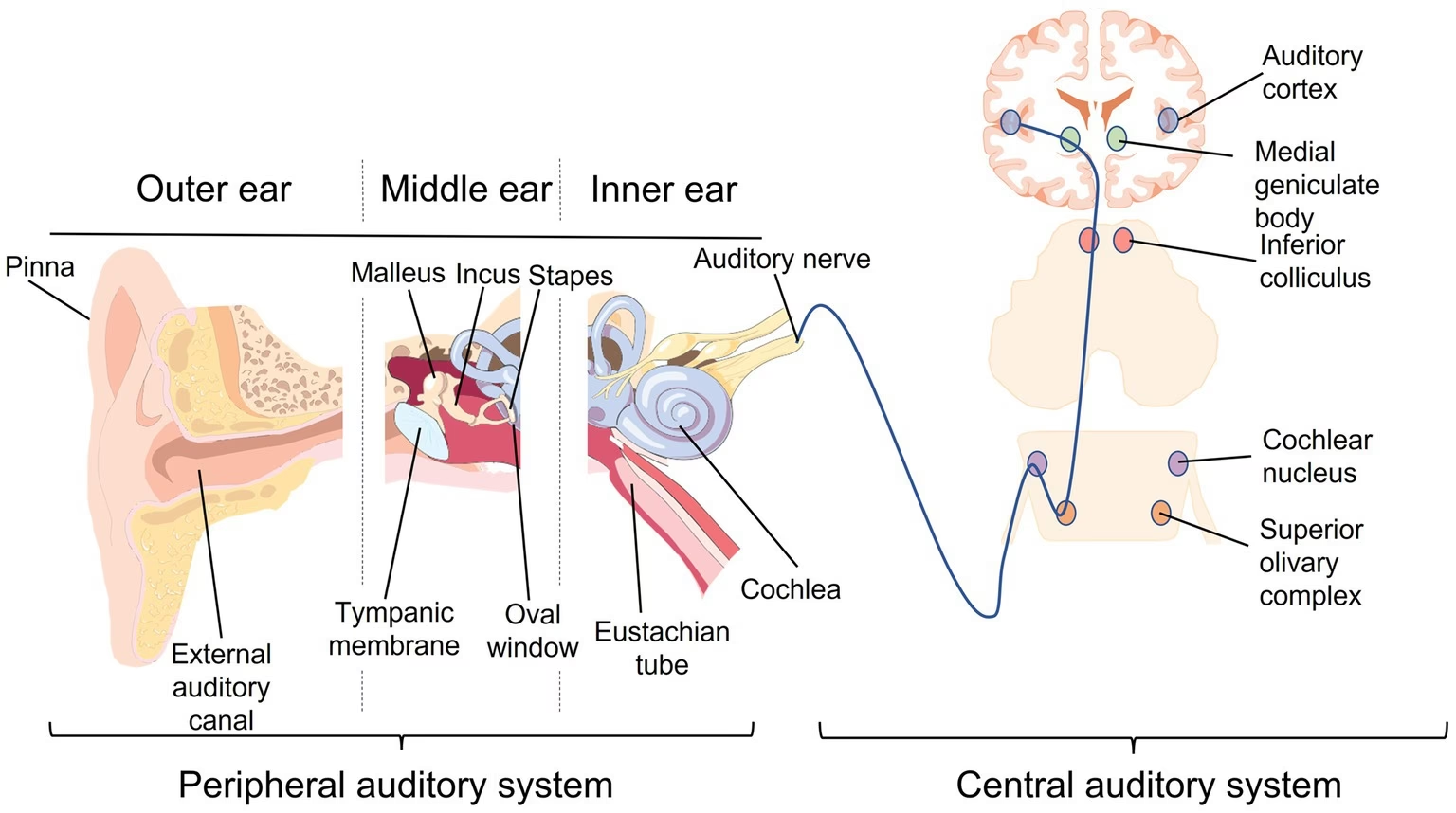
El córtex auditivo es la parte del córtex cerebral responsable del procesamiento de la información auditiva. Representa el punto donde terminan las fibras que ascienden desde el cuerpo geniculado medial (CGM) del tálamo, una estación de relevo crucial en la vía auditiva. En la vía auditiva ascendente, las diferencias entre especies son particularmente evidentes en el córtex auditivo. El número de áreas identificadas varía enormemente, desde tan solo cinco en ratones hasta más de 30 en algunos estudios detallados de humanos. Estas diferencias inter-específicas no solo se manifiestan en la cantidad de áreas, sino también en su posición relativa, disposición, densidad celular, patrones de conexión y la forma en que se organiza la representación de la frecuencia del sonido, conocida como tonotopía. Por ejemplo, mientras que en los humanos el córtex auditivo se localiza principalmente en el lóbulo temporal, en animales como el ratón se sitúa en la mitad caudal del córtex parietal.
Organización Compleja: Áreas Nucleares y de Cinturón
A pesar de la variabilidad entre especies, un principio organizativo común en el córtex auditivo de los mamíferos es la división en áreas "nucleares" (core) y áreas "secundarias" o de "cinturón" (belt). Las áreas nucleares suelen ser un koniocórtex (córtex granular) y contienen una o más representaciones completas y ordenadas de las frecuencias audibles. En estas áreas, como el córtex auditivo primario (AI) y el campo auditivo anterior (AAF) en la mayoría de los mamíferos estudiados, existe un cambio sistemático en la representación de frecuencia a lo largo de la superficie cortical. Las neuronas responden mejor a frecuencias específicas, organizadas de forma progresiva, como si fueran láminas apiladas de frecuencias bajas a altas. En el gato, por ejemplo, se han definido cuatro regiones con organización tonotópica: AI, AAF, campo auditivo posterior y campo auditivo ventroposterior.
Las áreas de cinturón rodean el núcleo y presentan una representación de frecuencia menos nítida o sonotopía, es decir, las neuronas no están tan finamente sintonizadas a frecuencias específicas y el cambio progresivo en la representación de frecuencia no es tan regular. En ratones, las áreas de cinturón incluyen el campo ultrasónico (UF), el campo auditivo secundario (AII) y el campo dorsoposterior (DP). Estas áreas carecen de una tonotopía clara y muestran propiedades de respuesta más complejas, como curvas de sintonización amplias, múltiples frecuencias óptimas y respuestas selectivas a modulaciones de frecuencia. La distinción anatómica entre las áreas de cinturón es a menudo menos clara que la de las áreas nucleares; generalmente, la densidad celular, la mielinización y la expresión de marcadores como la citocromo oxidasa o la parvalbumin están reducidas en comparación con el núcleo.
Arquitectura Celular en Capas: Una Estructura Laminar Sofisticada
El córtex auditivo exhibe la organización laminar característica del neocórtex, compuesto por seis capas (I a VI) con diferentes densidades y tipos celulares. Esta estructura en capas es fundamental para el procesamiento jerárquico y paralelo de la información auditiva.
- Capa I: La más externa, con la menor densidad neuronal. Más del 90% de sus células son pequeñas neuronas GABAérgicas.
- Capa II: Contiene neuronas piramidales y no piramidales. Las células más pequeñas están superficialmente, mientras que las piramidales más grandes se encuentran cerca del borde con la Capa III. Aproximadamente una cuarta parte de las no piramidales son GABAérgicas.
- Capa III: Poblada por varios tipos de neuronas piramidales y no piramidales. Las piramidales más grandes están en la parte profunda, cerca de la Capa IV. Sus dendritas apicales alcanzan la Capa I. Las no piramidales (estrelladas, multipolares) tienden a ser GABAérgicas.
- Capa IV: Principal destino de las proyecciones talámicas ascendentes. Compuesta principalmente por células estrelladas con somas y campos dendríticos orientados radialmente.
- Capa V: Contiene neuronas piramidales y no piramidales. Destacan las neuronas piramidales grandes y conspicuas, cuyas dendritas apicales se extienden hasta la Capa I. Esta capa es una fuente principal de proyecciones descendentes. Las células GABAérgicas son principalmente multipolares y bipolares.
- Capa VI: Presenta la mayor variedad de tipos celulares, incluyendo neuronas piramidales, multipolares, bipolares y horizontales. Menos del 20% son GABAérgicas. Es otra fuente crucial de proyecciones descendentes.
La distribución de marcadores como la Parvalbumin y la Calbindin varía entre las capas, reflejando la diversidad y organización de las poblaciones neuronales. Por ejemplo, la Parvalbumin muestra una inmunorreactividad intensa a lo largo de las capas II a VI.
La Intrincada Red de Conexiones del Córtex Auditivo
El córtex auditivo está integrado en vastas redes neuronales a través de diversas proyecciones:
- Proyecciones Ascendentes: La principal entrada excitatoria a las áreas nucleares (AI, AAF) proviene del CGM del tálamo, principalmente de la parte ventral (CGMv) y dorsal (CGMd), que terminan mayormente en la Capa IV. El CGM medial (CGMm) también proyecta al núcleo y de forma más difusa a todas las áreas corticales auditivas. La organización de estas conexiones a menudo refleja la tonotopía.
- Conexiones Intracorticales: Las áreas dentro del córtex auditivo ipsilateral están interconectadas, siendo las conexiones más densas entre sitios con representaciones de frecuencia similares.
- Conexiones Comisurales: El córtex auditivo de un hemisferio se conecta con el córtex auditivo contralateral, concentrándose en áreas homótopas (correspondientes) y vinculando fuertemente representaciones de frecuencia idénticas.
- Proyecciones Descendentes (Corticofugales): Las capas V y VI son las principales fuentes de proyecciones que salen del córtex auditivo. Estas proyecciones se dirigen a estructuras subcorticales (tálamo, mesencéfalo, puente) y al hemisferio contralateral a través del cuerpo calloso.
Un aspecto particularmente interesante de las proyecciones descendentes es la distinción funcional entre las neuronas corticotalámicas de la Capa V y la Capa VI. Las neuronas de la Capa V suelen ser piramidales grandes con dendritas apicales gruesas y a menudo muestran "descarga intrínseca en ráfagas" en respuesta a estímulos. Se consideran "impulsoras" (drivers), transmitiendo señales robustas y fiables a sus objetivos subcorticales. Por otro lado, las neuronas corticotalámicas de la Capa VI son más pequeñas, con dendritas apicales más cortas y delgadas, y generalmente presentan "descarga regular". Se les considera "moduladoras", integrando información localmente y ajustando la excitabilidad de sus objetivos subcorticales. Esta diferencia en sus propiedades eléctricas intrínsecas y en los patrones de entrada que reciben sugiere que las proyecciones corticotalámicas de la Capa V y VI desempeñan roles distintos en el procesamiento de la información, influyendo en cómo el tálamo y otras estructuras responden a las señales auditivas.
Especialización Funcional y el Procesamiento del Sonido Complejo
La división entre áreas nucleares y de cinturón no es solo anatómica, sino que también refleja una especialización funcional en el procesamiento auditivo. Las áreas nucleares, con su precisa tonotopía y neuronas finamente sintonizadas, son cruciales para el análisis de las características acústicas básicas, como la identificación de tonos puros y la discriminación fina de frecuencias e intensidades. Constituyen el "núcleo" del procesamiento auditivo, donde la información se organiza de manera fundamental.
Las áreas de cinturón, o áreas no lemniscales, son anatómicamente y fisiológicamente distintas del núcleo. Son generalmente más numerosas y, aunque menos estudiadas en detalle, se sabe que procesan aspectos más complejos del sonido. Sus neuronas responden a características como la modulación de frecuencia, el inicio o el cese de los sonidos, y pueden integrar información de diferentes frecuencias de manera menos rígida que las áreas nucleares. Crucialmente, las áreas de cinturón tienen conexiones más amplias con regiones fuera del córtex auditivo propiamente dicho, incluyendo áreas de los lóbulos temporal, frontal y parietal. Esta extensa conectividad sugiere que las áreas de cinturón son fundamentales para integrar la información auditiva con otras modalidades sensoriales, el contexto cognitivo y las funciones de orden superior, como la atención, la memoria y la toma de decisiones basadas en estímulos auditivos. Se cree que esta organización soporta la existencia de vías de procesamiento paralelas, donde el núcleo se encarga de la "qué" del sonido (identificación de características básicas) y las áreas de cinturón contribuyen al "dónde" (localización espacial) y al "qué significa" (reconocimiento de patrones, significado contextual).
Comparativa: Áreas Nucleares (Core) vs. Áreas de Cinturón (Belt)
| Característica | Áreas Nucleares (Core) | Áreas de Cinturón (Belt) |
|---|---|---|
| Tonotopía | Clara y ordenada | Menos definida o ausente |
| Sintonización Neuronal | Aguda (a frecuencias específicas) | Más amplia o compleja |
| Procesamiento Principal | Características acústicas básicas (frecuencia, intensidad) | Características complejas (modulación, patrones), integración multimodal |
| Conexiones Principales Ascendentes | CGMv y CGMd (proyecciones topográficas) | CGMd y CGMm (proyecciones más amplias) |
| Conexiones Extracorticales | Menos extensas | Más extensas (a áreas temporales, frontales, parietales) |
| Ejemplos (Ratón) | AI, AAF | UF, AII, DP |
Preguntas Frecuentes sobre el Córtex Auditivo
- ¿Cuál es la función principal del córtex auditivo?
- Su función principal es el procesamiento de alto nivel de la información sonora. Recibe señales del tálamo y las transforma en percepciones auditivas conscientes, permitiendo la identificación, discriminación y localización de sonidos complejos.
- ¿Qué significa que un área del córtex auditivo sea tonotópica?
- Implica que existe una organización espacial donde las neuronas están dispuestas según la frecuencia del sonido a la que responden de manera óptima, creando un mapa ordenado de frecuencias.
- ¿Cómo se diferencia el procesamiento en las áreas nucleares y de cinturón?
- Las áreas nucleares se centran en el análisis de características básicas como la frecuencia y la intensidad (con tonotopía clara). Las áreas de cinturón procesan características más complejas, integran información de diferentes frecuencias y se conectan con otras regiones cerebrales para contextualizar el sonido (con tonotopía menos definida o ausente).
- ¿Qué papel juegan las capas V y VI en las proyecciones del córtex auditivo?
- Estas capas son la fuente principal de proyecciones descendentes (corticofugales) que envían información de vuelta al tálamo y otras estructuras subcorticales. Las neuronas de la Capa V actúan como "impulsoras" de señales, mientras que las de la Capa VI actúan como "moduladoras", ajustando la excitabilidad de sus objetivos.
- ¿El córtex auditivo es igual en todos los mamíferos?
- No, existen diferencias significativas entre especies en cuanto al número de áreas, su organización espacial, densidad celular, conexiones y representación tonotópica. Sin embargo, la división general en áreas nucleares y de cinturón es un patrón común.
En resumen, el córtex auditivo es una estructura cerebral notablemente compleja y dinámica, esencial para nuestra capacidad de percibir, interpretar y responder al mundo sonoro que nos rodea. Su organización jerárquica y en paralelo, junto con su intrincada red de conexiones, le permite realizar desde el análisis más básico de las propiedades acústicas hasta la comprensión más elaborada de la información auditiva. La investigación continua de sus circuitos y mecanismos celulares sigue siendo clave para desentrañar completamente los misterios de la percepción sonora en el cerebro.
Si quieres conocer otros artículos parecidos a El Córtex Auditivo y el Procesamiento del Sonido puedes visitar la categoría Neurociencia.